El Mioceno es la cuarta división del
periodo terciario del cenozoico, que comenzó hace 23,3 millones de ańos y
finalizó hace 5,2 millones de ańos.
La elevación de las grandes cordilleras montańosas que había comenzado como
resultado de la colisión entre placas tectónicas durante el oligoceno, siguió
adelante. Entre estas cordilleras, las principales fueron los Alpes en Europa,
el Himalaya en Asia y las cadenas montańosas del continente americano. Los
sedimentos producidos por la erosión de estos sistemas se depositaron en cuencas
marinas poco profundas, para terminar convirtiéndose en la localización de ricos
depósitos petrolíferos en California, Rumania y la costa oeste del mar Caspio.
El clima del mioceno era más fresco que el de la época precedente. En el
hemisferio sur se había establecido ya un sistema circumplanetario de corrientes
oceánicas, que aislaba a la Antártida de las corrientes más cálidas del resto
del mundo.
Esto favoreció la
aparición de un gran casquete de hielo antártico. En el hemisferio
norte, grandes áreas antes cubiertas por espesos bosques se
convirtieron en grandes praderas. La fauna del mioceno incluía una
serie de mamíferos, entre ellos el rinoceronte, el camello, el gato
y el caballo. El mastodonte hizo también su aparición, al igual que
el mapache y la comadreja. Durante esta época, los grandes simios,
relacionados con el orangután, vivían en Asia y en la parte sur de
Europa; estos simios del mioceno son los parientes más próximos de
los simios humanoides que aparecieron en el plioceno.
América Central.
Los
monos del Nuevo Mundo (platirrinos) forman parte de los ecosistemas
tropicales modernos tanto del norte como del sur de América. Sin
embargo, estas dos gigantescas masas de tierra estaban separadas por un
océano hasta que el istmo de Panamá los unió hace unos 3,5 millones de
ańos. Estimaciones genéticas sugerían que los monos no llegaron a
América Central, la parte más meridional de la masa continental de
Norteamérica, hasta este momento, pero la ausencia completa de fósiles
había limitado el conocimiento de su historia. Los nuevos
fósiles del Panamacebus fueron encontrados en la formación Las
Cascadas gracias al trabajo de los investigadores, que se
vieron obligados a correr detrás de los ingenieros de las
obras cada vez que dinamitaban las empinadas orillas del
canal. Creemos que el Panamacebus estaba relacionado con el
capuchino (también conocido como mono cariblanco) y con los
monos ardilla que se encuentran en América Central y del Sur
en la actualidad.
Asia (Japón)
y América del Norte (costa del Pacífico).Era
un ungulados, y tenia 1.8 m de longitud Desmostylus era
un miembro típico de este grupo. Tenía la forma de un
hipopótamo y tal vez su comportamiento también fuera
similar; e! cuerpo era grueso y las patas robustas, con pies
anchos y cuatro dedos ungulados en cada uno. Los huesos de
la parte inferior de las patas estaban fusionados, formando
un pilar sólido, lo cual significaba que para girar el pie
tenía que dar la vuelta a toda la extremidad. Debajo del
agua, es probable que el animal se fuera impulsando, como
hacen los hipopótamos modernos cuando «caminan» sobre el
lecho de los ríos. La parte anterior de las mandíbulas,
tanto superiores como inferiores, era alargada y portaba una
serie de colmillos que apuntaban hacia adelante, formados
por los incisivos alargados y por los caninos. La cabeza del
animal debió de parecerse a la de los elefantes de colmillos
en forma de paleta, que vivían en esa época.
Desmostylusdebía de tener una forma de vida anfibia, impulsándose por
las aguas poco profundas cercanas a la costa y arrancando
moluscos de las rocas con sus colmillos
delanteros. También es posible que se haya zambullido hasta
e! fondo del mar en busca de alimentos. Algunos
paleontólogos suponen que en medio de las mareas se
alimentaba de algas marinas.
Perú. Una
expedición de investigadores peruanos encontró fósiles de un
cocodrilo de unos 14 metros, el mayor depredador que acechó
en la Amazonia, cuando en lugar de la espesa selva había un
gran mar tropical, hace 15 millones de ańos. Fueron
hallados restos del gigantesco esqueleto del cocodrilo, así
como la mandíbula y la dentadura, bajo el fango a unos 480
kilómetros al noreste de Lima. El cocodrilo pesaba unas
nueve toneladas y la cabeza tenía una longitud de 1.3
metros, mientras que el cuerpo tenía una envergadura de unos
14 metros. Este es el segundo fósil de este tamańo y
características que descubren los científicos en las tres
últimas décadas, según afirmó Rodolfo Salas, responsable del
área de paleontología del Museo de Historia Natural de Perú.
Los expertos creen que el cocodrilo se alimentaba de
tortugas gigantes, y en el museo de Lima ya hay al menos un
fósil de una tortuga de estas características con las marcas
de una mordedura del depredador. “El hallazgo del fósil del
Purussaurus ayudará a reconstruir su ambiente
y a recuperar así un poco del tiempo perdido en el pasado”,
dijo Salas. Cerca de los restos del cocodrilo los
científicos también encontraron fósiles de otros reptiles
más pequeńos, así como de perezosos y armadillos gigantes.
Bagua Chica, Perú.
Gracias a unos restos fósiles
hallados en Bagua, se ha podido confirmar la
existencia en el territorio peruano de esta terrible
bestia a quienes los científicos conocen como
piroterio.
El implacable sol causaba estragos sobre su espalda,
pero ni el calor ni el sudor lograban detener sus
manos que sujetaban el cincel y la picota de geólogo
con firmeza, como si vaticinaran que la brújula y su
antiguo localizador satelital (GPS) lo conducirían
al tesoro más codiciado entre sus colegas
paleontólogos. "Los geólogos a veces tenemos ojos
infrarrojos. Nos llama la atención cualquier piedra
y nos lanzamos a buscar", rememora y a las pruebas
se remite. Ante sus ojos y los del equipo que
colaboraba en la excavación, desarrollada en el
sector El Milagro, a diez minutos de la ciudad de
Bagua Chica, aparecieron el maxilar, algunas
vértebras y dientes de un piroterio, uno de los
mamíferos menos estudiados del continente, debido a
los escasos vestigios de su tipo descubiertos a la
fecha.
Con los restos desenterrados en Bagua, la comunidad
científica pudo finalmente estimar el tamańo del
animal, casi tres metros de largo y 2,5 toneladas de
peso. Igualmente se verificó que su morfología era
similar al 'Pyrotherium romeroi',
especie del que se conocía gran parte del esqueleto.
Pero ni siquiera son parientes. A diferencia del
romeroi, el piroterio de Bagua, 'Baguatherium',
era más pequeńo y primitivo. Para el hombre
contemporáneo parecería un híbrido entre rinoceronte
y sachavaca, con una pequeńa trompa, colmillos y
patas de elefante. Estos fósiles son de una época
muy especial en la evolución de la Tierra porque fue
entonces cuando se extinguieron los dinosaurios.
Hace 60 millones de ańos Sudamérica semejaba una
isla, alejada del resto de tierra firme formada por
lo que hoy se conoce como América del Norte, África,
Europa, Asia y Oceanía. Mientras animales como los
primitivos caballos y camélidos empezaban su
evolución en el hemisferio norte, un peculiar grupo
de mamíferos se diversificaba miles de kilómetros al
sur, durante el período de aislamiento geográfico
cuyos resultados aún concitan interés. Los huesos
fosilizados de estas especies, que se encuentran
entre rocas del período Terciario, pertenecen a
linajes ausentes en los demás continentes, razón por
la que cuentan con una historia evolutiva propia.
Siempre que se habla de los animales representativos
del Perú nos referimos a los camélidos, al oso
andino y al puma. Casi nadie sabe que estos animales
invadieron Sudamérica hace relativamente poco tiempo
y que los verdaderos mamíferos nativos de nuestro
territorio fueron animales totalmente distintos",
dijo. "El de Bagua fue el más grande que existió en
su época. Hasta hace 10 ańos sus fósiles eran
prácticamente desconocidos en territorio peruano.
Hubo hallazgos en Argentina, Brasil, Colombia y
Bolivia, pero limitados. Sin embargo, gracias a los
restos encontrados recientemente en nuestro país,
ahora se sabe que existieron diversas especies,
muchas de ellas nuevas para la ciencia. Decididos a
que la reconstrucción paleontológica peruana
abandone de a pocos su anonimato, cinco
especialistas del Museo de Historia Natural
elaboraron un modelo a escala real del piroterio de
Bagua, que esperan sea reconocido como 'Baguatherium
jáuregui' en homenaje al lugar donde se
descubrió y al funcionario del INC que colaboró con
las excavaciones.
Urumaco, Venezuela.
Corresponde a un género extinto de tortugas de agua
dulce. Sus fósiles han sido encontrados en el norte
de América del Sur, concretamente en Venezuela, en
rocas que datan de fines del Mioceno al inicio del
Plioceno, hace aproximadamente 6 a 5 millones de
ańos. De la cantidad de fósiles hallados en la
expedición de 1972 a Urumaco, se encuentra el
caparazón de 23 dm de largo y 19,5 dm de ancho, que
dio origen a una nueva especie bautizada con el
nombre científico de Stupendemys geographicusque proviene de la combinación de "estupenda",
cualidad atribuida a la especie, y el nombre de la
organización "National Geographic". En Venezuela se
conoce a esta especie bajo el nombre afectivo de
uruma, tal como la bautizó el Museo de Ciencias de
Venezuela. Según el Dr. Roger Wood, la uruma es el
género y la especie de tortuga más grande que haya
existido sobre la faz de la Tierra y se cree que se
extinguió hace aproximadamente 6 millones de ańos
atrás. De los estudios realizados al fósil, se pudo
conocer que se trataba de una tortuga acuática y
seguramente ovípara. Su alimentación debió ser de
material vegetal y de invertebrados acuáticos. De
las tortugas actuales, la Arrau es hasta el
momento la más parecida a Uruma, siendo la
más grande de las aguas continentales de Venezuela,
llegando a medir el especímen hembra hasta 9 dm y 5
dm el macho.
Región de Ica, Perú y
Argentina.
Es
un género y especie extinto de odontoceto, perteneciente a
la familia Physeteridae conocido por un único ejemplar que
habitó en lo que ahora es Perú. Se encontró en el desierto
de Pisco, Región de Ica, en rocas de hace 12-13 millones de
ańos. Se han encontrado restos fósiles del 75% del cráneo y
grandes fragmentos de los maxilares, así como varios dientes
(los mayores medían 36 cm de longitud, mucho más que el
actual cachalote). Su cráneo medía unos 3 metros de largo y
la longitud total debió ser de 13,5 a 17,5 metros. La
ballena más parecida a Livyatan melvilleies
el actual cachalote (Physeter macrocephalus),
aunque la diferencia consiste en que Livyatan poseía
dientes en ambos maxilares. Se supone que era un predador
agresivo y posiblemente cohabitó y tal vez luchó con otro
superdepredador, Carcharodon megalodon, un
tiburón que pudo llegar a medir hasta 16 metros de longitud.[
Restos fósiles de muchos animales
como ballenas barbadas, zifios, delfines, marsopas,
tiburones, tortugas marinas, focas y aves marinas se
hallaron en el mismo sitio donde los fósiles deL.
melvilleise extrajeron.L. melvilleipudo
haber sido el superdepredador de su tiempo, junto con el
tiburón gigante, Megalodon, contemporáneo de
L. melvilleien el mismo hábitat, y
probablemente tenía gran impacto en la estructura de las
comunidades marinas del Mioceno. La aparición de cachalotes
depredadores en el registro fósil coincide con una fase de
diversificación e incremento del tamańo y rango de
distribución de los misticetos (ballenas barbadas) en el
Mioceno.
Parángula, Venezuela.
Pertenece a un grupo extinto de reptiles crocodiliformes
venezolano clasificado en la Infraorden Sebecosuchia,
descubierto en la Formación de Parángula en el estado
Barinas. Su nombre deriva del término “Barinas”,
denominación del estado en Venezuela al que pertenece la
localidad donde fue hallado y de la latinización en
masculino del término griego “souchos”, que significa
cocodrilo. El cráneo de 70 cm que comparativamente con otras
especies su largo total pudiera llegar hasta 6 m de
longitud. Era carnívoro y carrońero. Vivió en el Mioceno
medio entre 13 y 15 millones de ańos, en Sudamérica. Pudo
haber alcanzado una velocidad aproximada a los 20 km/h a
cuatro patas y un poco más rápido si lograba una marcha
bípeda en cortos trayectos. El Barinasuchus arveloi tenia
narinas que se ubican en la punta de la cabeza, lo que
sugiere que pudo haber vivido más en tierra que en el agua,
hay que recordar que las narinas de los crocodiliformes
acuáticos actuales normalmente se encuentran de lado a lado
sobre una prominencia en la cima del hocico, para respirar
sin tener que exponer toda la cabeza fuera del agua y de esa
manera emboscar a sus presas en el momento que necesiten el
preciado líquido. Motivado a todas las características
anteriormente descritas, se conjetura que el Barinasuchus
arveloi debió haber habitado en bosques tropicales y
subtropicales.
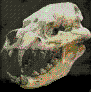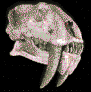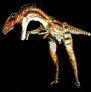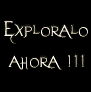
Cráneo en
el
Museo Jacobacci,
San Antonio Oeste, Rió Negro aportado por la Fundación Azara.
Cráneo exhibido en el Museo
Argentino de Ciencias Naturales de Buenos Aires
Reconstrucción en vivo (*)
Argentina.Durante
mucho tiempo, los carnívoros más exitosos en América del sur fueron
aves gigantescas como los Phorusrhacos. Es un género de gigantescas
aves depredadoras no voladoras que habitaron la Patagonia, de la
familia de las Phorusrhacidae, conteniendo una sola especie; el Phorusrhacos longissimus. Sus parientes más cercanos en
la actualidad son los de la familia Cariamidae, llamados también
Seriemas o Chuńas, pero su tamańo era mucho mayor que el de éstos;
siendo su aspecto más parecido al de un avestruz. Estas aves
habitaron en bosques y pastizales. Entre los huesos hallados en el
estrato de la Formación Santa Cruz (ahora considerado como
principalmente de mediados del Mioceno fecha) fue la pieza de una
mandíbula que Florentino Ameghino descubierto a principios de 1887 y
el mismo ańo en que describió por primera vez como una de
desdentados mamífero que él nombró Phorusrhacos longissimus.
El nombre específico significa "muy largo" en latín , de nuevo en
referencia a las mandíbulas inferiores. El holotipo es el ejemplar
mandíbula, MLP-118 (Museo de La Plata). En 1889 Ameghino corrigió el
nombre a una más correcta gramaticalmente Phororhacos pero el nombre
más temprana tiene prioridad. En 1891, fue reconocido por él como un
pájaro. Los restos se conocen de varias localidades de la provincia
de Santa Cruz , Argentina. Phorusrhacos situó en torno a 2,5 metros
de alto y pesaba unos 130 kilos. Fue apodado el "Terror Bird" por
razones obvias: es uno de los mayores carnívoros aves que jamás haya
existido, y sus alas rudimentarias formadas como estructuras de
brazo con garras en forma de un gancho de carnicero para presa
frente, que fue asesinado luego con el pico enorme. El ave mataba de
dos maneras. En primer lugar sujetando a su presa con su pico y
romper contra el suelo varias veces al igual que sus parientes
modernos, los chuńas. En segundo lugar, y más espectacularmente, al
golpear hacia abajo con la punta hacia abajo agudo en la parte
delantera de su pico superior. Si en la posición correcta, la serie
especulado, la espiga podría penetrar en el cráneo y matar
instantáneamente. Tenía un cráneo enorme de hasta sesenta
centímetros de largo, armado con un poderoso gancho con punta de
pico. La estructura del pico y las garras grandes en los dedos
demostrar que se trataba de un carnívoro.
Kelenken
guillermoi.Bertelli et al., 2007.
Esqueleto de
Kelenken recreado por la Fundación Azara a partir de
los restos conocidos.
Cráneo de
Kelenken guillermoi, descubierto en Comallo, Provincia de Rió
Negro, mide 70 centímetros de largo.
(*)
Posible
aspecto de
Kelenken guillermoi
(*).
Argentina.
Es una especie extinta de ave gruiforme de la
familia Phorusrhacidae o aves del terror. Es el ave predadora
conocida más grande de la historia. Vivió en el Mioceno, hace unos
15 millones de ańos. Midió más de tres metros de altura, su cabeza
(incluyendo el pico) llegó a medir más de 70 cm de longitud, siendo
el ave conocida con el cráneo más grande, llegaban a pesar los 160
kilogramos. Kelenkenera ágil y podía cazar y devorar
roedores, reptiles y pequeńos mamíferos; las crías de estas aves
alcanzaban los 100 km/h pero los adultos sólo llegaban a los 60 km/h.
Su fósil fue encontrado en la Patagonia, concretamente en la
estación de tren de Comallo, cerca de la ciudad de Bariloche,
Argentina por Guillermo Oscar Aguirrezabala. Los restos constan de
un cráneo y fragmentos de un pie y de una pata y se encuentran en el
"Museo Paleontológico de Bariloche. Antes del descubrimiento de su
cráneo, los científicos interpretaban las cabezas de las grandes
aves del Cenozoico como versiones a mayor escala de las de las aves
de menor tamańo. Los huesos de la pata y el pie tienen un parecido
superficial con los del ńandú, un ave no voladora actual de América
del Sur.
Argentavis
magnificens.Campbell & Tonni,
1980.
Cráneo de Argentavis
magnificens. Foto gentileza de Paleogenesis.
El Paleontólogo
Rosendo Pascual, comparando el humero de Argentavis y un
cóndor. (Discovery Channel).
Argentavis magnificens
atacando a un Thylacosmilus atrox. (*)
Mariano Magnussen Saffer
del Museo Municipal Punta Hermengo de Miramar, junto al ala
de Argentavis magnificens, recreada para esta
institución a partir de los pocos restos conocidos.
Argentina.Un
esqueleto parcial de este enorme teratornítido se encontró de la
provincia de La Pampa, Argentina. Es la mayor ave voladora que haya
existido. Es también el teratornítido más antiguo encontrado, ya que
data de fines del Mioceno, entre 6 y 8 millones de ańos, y uno de
los escasos teratornítidos encontrados en América del Sur. El primer
descubrimiento incluyó porciones del cráneo, un húmero incompleto y
varios otros huesos del ala. Incluso los cálculos conservadores
estiman su envergadura en por lo menos 6 metros, y podría haber
llegado hasta los 8 metros. Su peso se estima en alrededor de 80 kg.
Su peso y su envergadura rivalizan entonces con los de los más
grandes pterosaurios. Por su tamańo, seguro que fue un ave
planeadora, limitando el aleteo al carreteo y posterior despegue,
aprovechando las fuertes corrientes térmicas para su majestuoso
vuelo que provenían del océano Pacifico, antes de que las
cordilleras llegaran a las alturas que presentan en la actualidad. Por lo tanto,
Argentavis magnificens resulto ser hasta el momento el
ave voladora de mayor tamańo del mundo.
Gryposuchus
neogaeus. Burmeister, 1885.
Cráneo exhibido en el Museo
de La Plata.
Paleoreconstruccion. (*)
Argentina y Venuzuela. Es un género extinto
de crocodiliano gavialoide. Es el género tipo de la subfamilia
Gryposuchinae. Sus fósiles han sido hallados en varios países de
Suramérica, como Argentina, Colombia, Venezuela, Brasil y Perú. El
género existió durante principios y mediados del período Mioceno.
Una especie recientemente descrita, G. croizati, tiene
una longitud estimada de 10 metros. La especie tipo de
Gryposuchus esG. neogaeus. Especimenes de
esta especie fueron descritos inicialmente de Argentina en 1885,
aunque fueron referidos al género Ramphostoma.No fue
sino hasta 1912 que se le asignó su propio género con la descripción
de una nueva especie de Brasil, G. jessei. Algunos
gaviales griposuquinos como Siquisiquesuchus y
Piscogavialis han sido hallados en localidades
que se cree eran depósitos de ambientes costeros. La presencia de Gryposuchusen la formación Urumaco de Venezuela, que
incluye estratos marinos, le da crédito a la idea de que los
griposuquinos pueden haber vivido en ambientes costeros. Sin
embargo, ciertas localidades donde se recuperó material
perteneciente a la especie G. colombianus, como la
zona de La Venta, Colombia , claramente eran depósitos de ambientes
de agua dulce, lo que indica que la hipótesis de un estilo de vida
costero para los griposuquinos no puede generalizarse.
Boidae.
Gray, 1825.
Argentina. Los boidos o boas son
una familia de serpientes constrictoras, es decir que matan a sus
presas por constricción, encerrándolas en sus anillos. A diferencia
de sus cercanas parientes, las pitones (Pythonidae), son
ovovivíparas. A ambos lados de la cloaca presentan espolones, que
son vestigios de las patas posteriores locomotoras. Entre las boas
se encuentran algunas de las especies de serpientes más grandes.
Pero en el actual desierto patagónico, hace unos 15 millones de ańos
existía una exuberante selva, semejante a la amazónica, con grandes
lagartos, perezosos, monos y mucho más.
Hace algunos ańos atrás, un Paleontólogo
del Museo Argentino de Ciencias Naturales de Buenos Aires se
encontraba revisando las colecciones de dicha institución, hallo el
fragmento de una enorme vértebra que, en un principio, fue
atribuida a restos de un Dinosaurio. Al consultar los registros del
inventario del Departamento de Paleontología de Vertebrados
"Florentino Ameghino" del Museo, se dieron cuenta los
investigadores de que se trataba de un enorme vertebrado que vivió
durante el Terciario, mucho después de la desaparición de los
grandes reptiles de la Era Mesozoica. La pieza en cuestión fue
hallada por el Geólogo Roberto Ferello en el ańo 1953, en
sedimentos de origen continental de la zona comprendida entre los
lagos Musters y Colhue Huapi, al sur de la Provincia de Chubut. Hoy
sabemos que esta vértebra pertenece al Ofidio (Serpientes y víboras)
mas grande de todos los tiempos. Calculamos después de hacer varias
comparaciones con especies vivientes y extinguidas, este enorme
animal midió unos 20 metros de largo. Su cráneo tuve que medir
unos 70 centímetros, con lo que la abertura bucal le permitía
engullir presas de mas de un metro de circunferencia, como losAstrapotheriosy otros Notoungulados de la época.
Borhyaena
macrodonta.
Ameghino, 1897.
proborhyaena proborhyaena
proborhyaena proborhyaena
Esqueleto de borienido, en el
MEF.
Cráneo y mandíbula en el Museo
de La Plata.
Aspecto. Tomado de Prehistoric
Fauna.
Argentina.
Era parecido a un oso, con las patas planas. Fue un marsupial carnívoro
del orden Sparassodonta
del tamańo de un lobo, de 1,5 metros de longitud. Eran depredadores
muy importantes durante el Mioceno de la patagonia Argentina, los
cuales se alimentaba de mamíferos notoungulados y armadillos. Las patas cortas
seńalan que no era un animal corredor, pero seguro que asechaba a
sus victimas en los antiguos bosques que existían hace 20 millones
de ańos en los actuales desiertos de la provincia de Chubut.
Patagosmilus goini.
Forasiepi y Carlini, 2010.
Fragmento de cráneo con
molares y canino de Patagosmilusgoini. Colección Museo de La
Plata. (*)
Aspecto de
Patagosmilusgoini,
cazando unHuayqueriana
(*).
Argentina.Es un género
extinto de mamífero carnívoro metaterio de la familia
Thylacosmilidae que vivió a mediados del Mioceno en América del Sur.
Al igual que otros representantes de esta familia, como
Thylacosmilus atrox y Anachlysictis gracilis,
se caracterizaba por sus enormes
colmillos en la
mandíbula superior, similares a los conocidos felinos "dientes de
sable" (Machairodontinae), de los que eran equivalentes ecológicos
sin parentesco algunos. La morfología general dePatagosmilus
sugiere que era anatómicamente menos especializado que
Thylacosmilus del Plioceno, pero la morfología de los
dientes, sin embargo, sugiere que probablemente estaba más
estrechamente relacionado con este último, que con el más primitivo
Anachlysictis de Colombia. La especie
Patagosmilusgoini, que debe su nombre al
paleontólogo argentino Francisco Goín, especialista en marsupiales,
y fue descrita a partir de los restos de un cráneo aplastado y parte
de una falange ungual que se descubrió en sedimentos que datan del
Mioceno Medio en la orilla oeste del Río Chico, en la provincia de
Río Negro en la Patagonia, Argentina y depositados en el Museo de La
Plata. Este es el primer representante de Thylacosmilidae del que se
han encontrado restos en la Patagonia, y el primer género reconocido
aparte de Thylacosmilusy Anachlysictis
que es un miembro indiscutido de esta familia. Seguramente un un
importante marsupial depredador.
Astrapotherium
magnum. Owen 1853.
Astrapotherium magnun Astrapotherium
Astrapotherium magnun Astrapotherium magnun
Astrapotherium
Cráneo y
mandíbula de Astrapotherium magnun en el Museo Egidio
Feruglio (MEF).
Reconstrucción
en vivo del extrańo Astrapotherium magnun. Imagen Sergey
Krasovskiy.
Mandíbula
inferior. Colección Museo de La Plata.
Argentina. Mientras
los Marsupiales evolucionaban, los Notoungulados hacían lo mismo
para colonizar todos los ambientes.
Astrapotherium es un
género
extinto de
mamíferos
placentarios del
orden
Astrapotheria,
superorden
Meridiungulata propio de
Sudamérica. Eran del tamańo de un
rinoceronte y probablemente tenían una
trompa corta y flexible. Su nombre significa "animal estrella" o
"bestia del rayo".
El astrapoterio mejor conocido es Astrapotherium magnumdel periodo
Mioceno Inferior-Medio, cuyos restos provienen de las rocas
denominadas Formación Santa Cruz en
Argentina, que tienen unos 15 millones de ańos. Gracias a que
estaban atrapados en cenizas volcánicas, ha sido posible reconstruir
varios esqueletos completos de astrapotéridos. Un esqueleto casi
completo de este animal se encuentra en el Museo MEF de Trelew. La preservación de la columna vertebral casi completa y
articulada, hizo imposible extraerla con seguridad. De este modo el
ejemplar fue montado con las extremidades flexionadas, como si
estuviera echado sobre su vientre. El esqueleto permite apreciar que
los astrapoterios eran criaturas bastante distintas a ninguna otra.
Astrapotheriumera un animal grande, quizás tanto como un
búfalo africano (800 Kg) o un
rinoceronte indio (1000 Kg), su cuerpo era largo, de cerca de
2,7 metros de longitud y una altura a la cruz de 1,40 metros, con
patas relativamente cortas, muy similar en proporciones a un
hipopótamo. El
cráneo
de Astrapotheriumera corto pero la
mandíbula inferior era larga, los
caninos eran largos y los
molares macizos; los molares de los astrapoterios recuerdan
vagamente a los de los rinocerontes.
Cráneo y
mandíbula de Diadiaphorus del Mioceno de Chubut. Al lado,
detalle de la estructura de la extremidad. (*)
Aspecto de equino que
presentaba el extinto Diadiaphorus, un notoungulado extinto
de Sudamérica. (*)
Argentina.
La difusión de las planicies abiertas por todo el continente
sudamericano contribuyo a la evolución de unos animales corredores
de estructura ligera. Los proteroterios (primeras criaturas) eran
animales con aspecto de caballos, que vivieron entre el final del
Paleoceno hasta el final del Plioceno. Estos han experimentado
cambios evolutivos y adaptaciones paralelas a los verdaderos
caballos de Norteamérica, incluso mucho antes que en cualquier otro
sitio. El Diadiaphorus debió tener un aspecto muy
semejante al antílope de cuello corto o un poney. Tenia el tamańo
semejante al de una oveja, pero con las patas de un caballo de tres
dedos. Aunque los huesos pares de las extremidades (cubito, radio y
tibia, peroné) nunca llegaron a fundirse, como ocurrió después con
los caballos auténticos, las patas eran largas y delgadas. El digito
central (el tercero) era muy grande y soportaba todo el peso del
animal, mientras que los dos situados en cada lado (el segundo y el
cuarto) se habían atrofiado. La cabeza era corta y gruesa, y la caja
craneal era bastante grande. Debido a que las coronas dentarias eran
bajas, es probable que Diadiaphorus ramoneara la
vegetación mas blanda, los arbustos y los árboles de las planicies
patagónicas. Otra especie conocida; Diadiaphorus
caniadensis(Kramarz & Bond, 2005).
Oxyodontherium zeballozi.Ameghino 1883.
Cráneo de
Oxyodontherium zeballosi, exhibido en el Museo de Cs
Naturales y Antropológicas de Mendoza.
Aspecto de un
macrauquénido, por el paleoartista Daniel Boh.
Argentina. Era un animal más
robusto y corpulento que un caballo, de miembros largos, con tres
dedos en cada pie, como los tapires, un cuello largo como el de una
jirafa, cuyo nicho ecológico ocupaba con formas intermedias que
recuerdan al okapi y un cráneo pequeńo y provisto, en vida, con una
corta trompa parecida a la de un tapir. Estos macrauquénidos son una
familia extinta de mamíferos placentarios del orden Litopterna
perteneciente al superorden Meridiungulata. La dentadura de todo el
grupo presenta la dotación de los primeros mamíferos, con 44
dientes. El nicho ecológico fue posteriormente ocupado por
litopternos de mayor tamańo y en la actualidad en parte por el
guanaco.
Cráneo del
primate Homunculus in situ en el sedimento. (*)
Cráneo de
Homunculus patagonicus depositado en el Museo de
Paleontología de Córdoba.
Reconstrucción
en vivo. (*)
Argentina.
Es la única
especie del
género monotípico
extinto de
primate
platirrino Homunculus, el cual vivió en la
Patagonia argentina, en el sur de
América del Sur, durante el
Mioceno inferior.
Fue un primitivo primate (monos) aloctono, cuyos ingreso al
continente fue hace 30 millones de ańos según la evidencia fósil
reunida hasta el momento. Vivió en el Rió Pinturas, en la Patagonia Argentina durante la edad Santacrucense. El hallazgo de
estos mamíferos indican que durante parte del Mioceno temprano, el
sur Argentino tenia condiciones ambientales muy semejantes a las que
hoy podemos observar en la selva misionera y amazónica. Su tamańo
es muy pequeńo y no pesarían mas de 1 kilogramo. Al igual que sus
representantes vivientes, su dieta era omnívora. Comían insectos,
frutos, pequeńos mamíferos etc. Lo que mas llama la atención de
estos primates es su "encefalizacion progresiva", es
decir, el aumento de la corteza cerebral a comparación de su tamańo,
lo que le permitiría retener información e inteligencia, pero en
este periodo algo muy remoto. La especie tenía el rostro
relativamente corto y órbitas de moderado tamańo, lo cual sugiere
que se trataba de una especie con hábitos
diurnos. El cráneo era grácil y carecía de cresta sagital. Tenía
una masa corporal de alrededor de 2,7 kilogramos. Basándose en la
morfología dental, se trataba de una especie
frugívora que complementaba con hojas para suplir su dieta. De
acuerdo a los restos
postcraneales probablamente se trataba de un
cuadrúpedo. sinonimo Anthropops perfectus.
Glyptatelus tatusinus.
Ameghino, 1897.
Fragmentos de placas óseas
de Glyptatelus.
(*)
Aspecto en vida de
Glyptatelus por Julio Lacerda. (*).
Argentina y Uruguay.
Pertenecen
a una familia muy particular de Xenarthros extinguidos, los
"Glyptodontidae". Tenían una coraza ósea (Cingulata) autóctonos
de América del sur, de tamańo mediano, alcanzando una notable
diversificación durante el Plioceno y Pleistoceno. Su caparazón óseo,
espeso y rígido, formado por placas soldadas entre si, con dentición
especializada para la dieta herbívora, alimentándose
principalmente del pastoreo se convirtieron rápidamente en animales
exitosos para los pronunciados cambios climáticos que se
aproximaban. Su coraza no les permitía realizar grandes movimientos
ni realizar cuevas, ya que estas no presentaban bandas móviles como
los peludos y mulitas actuales. Su cola probablemente estaría
provista de unos anillos óseos y un tuvo largo al final de la
misma, como los ejemplares mas recientes extintos, pero la ausencia
por ahora de estos restos no afirma de que sea así. Su cráneo era
grande y tosco, cubierto en la parte superior por un escudete de
placas, lo que protegía al animal cuando era atacado por predadores
de la época, como las gigantescas Aves o los Marsupiales dientes de
sable. Sus patas eran muy cortas y robustas como para soportar el
peso del animal. Su fémur totalmente desproporcionado a comparación
a otros huesos largos, y la pelvis soldada al caparazón del animal.
El mismo vivió durante el Mioceno de Chubut, Argentina.
Pelecyodon cristatus. F. Ameghino,
1891.
Cráneo y
mandíbula de Pelecyodon cristatus, hallado en el extremo
austral de Patagonia, Estrecho de Magrallanes. Museo de
Paleontología de la Universidad de Córdoba.
Cráneo completo y mandíbula inferior de
Pelecyodon del Mioceno
temprano de Santa Cruz. Museo de La Plata.
Aspecto en vida
de Pelecyodon cuando la Patagonia era una selva tropical. (*)
Argentina.
Durante el Mioceno, hace 20 millones de ańos, los Xenarthros estaban representados y adaptados en América del sur, el cual,
en el Mioceno se encontraba geográficamente bien distribuido por la
actual patagonia Argentina. Pelecyodon era de tamańo chico, media hasta un
metro de largo, a diferencia de sus gigantescos parientes del
Pleistoceno que superaban los cuatro metros de altura. Tenia garras
muy desarrolladas y curvas (una característica común entre los mamíferos edentados) seguro debe haberse visto desplazarse sobre sus nudillos
anteriores. Es muy probable que pasara la mayor parte de su tiempo
en las copas de los árboles, sujetándose de sus garras y alimentándose
de frutos suculentos en los tupidos bosques sub tropicales del
Mioceno. Como todos los edentados, Pelecyodon tenia pocos dientes y
sin esmalte. Sus mandíbulas solo tenían cuatro pares de dientes. Si
bien hoy en día la Patagonia Argentina es un gran desierto, durante
el Mioceno estaba constituida por grandes y extensas selvas,
evidenciado por la diversidad faunistica y botánica recuperada. Este
animal esta emparentado con el Perezoso de Tres Dedos (Bradypus),
que vive actualmente desde el norte de Argentina a Nicaragua. Otras
generos relacionados; Eucholoeops ingens y
Pseudhapalops s.p.
Nematherium angulatum.
Ameghino, 1887.
Cráneo de
Nematheriumsp en el Royal Ontario Museum. (*)
Aspecto en
vida de Nematherium. (*)
Argentina.
Los pilosos son un orden de mamíferos placentarios
que incluye los osos hormigueros, los tamandúas y los perezosos.
En la actualidad existen únicamente en el continente americano. El
origen del orden se remonta a principios del Terciario (hace
alrededor de 60 millones de ańos, poco tiempo después de la
extinción de los dinosaurios). El género
Nematheriumde tamańo algo mayor a otros xenarthros,
unos 2,5 metros de largo, aunque la ausencia de fósiles, no ayuda a
tener la suficiente información. Serían de ambientes cerrados,
consumidores de hojas, frutos y posiblemente tubérculos o raíces
gracias a su mayor capacidad cavadora. Probablemente era de
movimientos lentos, usando sus garras delanteras como arma
defensiva. Aunque se consideró vegetariano, algunos científicos
plantean la posibilidad de que hubiese consumido alternativamente
carrońa., tema aun en discusión. Posiblemente al igual que otros
perezosos, exhibe la interesante característica de poseer pequeńos
osteodermos, huesos insertos en la piel, que presumiblemente
ańadirían alguna protección al animal. Sus restos fueron recuperados
en Formación Santa Cruz, Mioceno temprano de la Patagonia Argentina.
Otras géneros relacionados;Analcitherium
Ameghino (Mylodontidae) y Prepotherium
Ameghino (Megatheriidae).
Phoberomys burmeisteri.
Ameghino, 1886.
Fragmento de rama
mandibular de
Phoberomys en el Museo de La Plata.
Cráneo de Phoberomysrecreado en
cartón con interesantes detalles, realizado por Luciano
Reuil.
Tamańos comparativos
con un humano y un carpincho, el roedor viviente de
mayor dimensión. (*)
Argentina, Colombia y Venezuela.
Era un roedor caviamorfo endémico y
autoctono (?) de América del sur, de tamańo grande. Generalmente
cuando hablamos de roedores nos imaginamos las ratas o lauchas.
Phoberomys burmeisteri era algo mas grande que un cerdo domestico y superaría
los 200 kilos de peso. Esta emparentado con el súper roedor del
Plioceno "Telicomys" (ver). Su cuerpo era grande y de
aspecto rechoncho. Sus extremidades eran largas y angostas. Su
maxilares y ramas mandibulares estaban provistas por cuatro molares
y incisivos con 2 centímetros de espesor. Su dieta estaría
adaptada a los follajes mas duros de la región. Estos ejemplares
son solo conocidos por piezas aisladas que no aportan mucha
información sobre su morfología.
Palaeospheniscus bergi.
Moreno y Mercerat, 1891
Aspecto de como seria el esqueleto del pingüino mioceno
Palaeospheniscus.
(*).
Cráneo de
Palaeospheniscus comparado con
un pinguino actual de tamańo promedio (*).
Aspecto de
Sphenisciforme en estudio.
Imagen prensa.
Argentina y Peru.
El género Palaeospheniscus fue nombrado
por François Moreno y Alcide Mercerat en 1891 para designar una
serie de especímenes fósiles procedentes de la Patagonia argentina.
La especie tipo del género corresponde a Palaeospheniscus
patagonicus, cuyo holotipo consiste en un tarsometatarso
aislado que se encuentra depositado en el Museo de La Plata,
Argentina y proceden de la Formación Gaiman (Mioceno temprano).
Capas portadoras de fósiles marinos del Cenozoico
medio afloran en cerro Plataforma en la Provincia del Chubut. Aquí
fueron hallados restos de pingüinos y un diente de tiburón bien
preservados, además se encontraron fósiles de invertebrados se
incluyen corales, briozoos, abundantes moluscos, equinoideos y
cangrejos.
Muchos de los fósiles de pingüinos hallados en
Patagonia han sido asociados a este género, al punto que en 1946
George Gaylord Simpson mencionaba nueve especies (la mayoría de las
cuales Simpson consideraba sinónimos de la especie tipo). El mismo
autor, en una revisión posterior realizada en 1975, solo consideraba
validas cuatro especies. En la actualidad solo se incluyen tres
especies en el género: Palaeospheniscus patagonicus,
Palaeospheniscus bergi y Palaeospheniscus
biloculata. Igualmente, los géneros patagónicos
Paraspheniscus, Trelewdyptes,
Chubutodyptesy Perispheniscus han sido
actualmente incluidos en alguna de estas tres especies. Los análisis
recientes sugieren que Palaeospheniscus se encuentra
más próximo a los pingüinos vivientes que a los pingüinos gigantes
del Paleógeno o al género Paraptenodytes. Seguramente
fue un gran nadador y se alimentaba principalmente de peces. Llego a
medir unos 80 centímetros de altura.
Califórnia, Texas,
Estados Unidos.Uno de los
últimos en la gran línea de evolución de los caballos fue el
Pliohippus, un caballo que se extinguió hace
unos cinco millones de ańos. El Pliohippus
evolucionó en América del Norte hace unos 15 millones de
ańos. Fue el primer caballo en tener un solo dedo en cada
pata. Los caballos más primitivos sólo tocaban el suelo con
un dedo, pero aún tenían dos dedos laterales en forma de
pequeńas sobrecańas. El Pliohippus era
ligeramente más pequeńo que los caballos modernos, y sus
dientes no estaban tan profundamente enraizados. Es una
forma importante puesto que se extendió por América del
Norte y dio lugar a una forma que invadió América del Sur (y
que ya se extinguió) y al verdadero caballo moderno que se
extendió por el mundo y fue domesticado más tarde por los
humanos.
Miohippus validus.
Osborn 1894.
Estados Unidos.Es un
género extinto de équidos de la subfamilia Anchitheriinae.
Los fósiles se encuentran en Norteamérica; parece haber
evolucionado del género Mesohippus, con el que
coexistió durante cuatro a ocho millones de ańos. Los
ejemplares pesarían entre 40 y 55 Kg, siendo más grandes que
la mayoría de los primeros équidos del Eoceno, pero menores
que los caballos actuales. Era algo más grande que
Mesohippus, y con el cráneo ligeramente más largo.
También tenía una cresta adicional en sus molares
superiores, lo que le permitía masticar hierba con mayor
efectividad. Algunas especies estaban adaptadas a la vida en
los bosques, y otras a la vida en las praderas. Miohippus
fue nombrado por Marsh en 1874 y su especie tipo es
Miohippus annectens. Fue asignado a la familia
Equidae por Marsh en 1874. Fue sinonimizado subjetivamente
conMesohippuspor Matthew en 1899.
Califórnia, Estados Unidos.
Las investigaciones que se han realizado gracias a este
nuevo fósil, revelan que este animal era un diestro
depredador de otros animales marinos gracias a sus robustas
mandíbulas con grandes caninos. Además, este tipo de morsa,
conocida como Pelagiarctos
y que ya se extinguió, era capaz, gracias a su morfología,
de devorar grandes presas sin discriminar: se alimentaba de
peces, invertebrados y, en ocasiones, podía también comer
presas de sangre caliente. Lo más característico de las
mandíbulas de esta morsa asesina es que son muy grandes y
sus huesos se fusionan en la barbilla, como en el caso de
los hombres. Los grandes dientes son parecidos a los que
tienen los animales 'rompe-huesos' como las hienas. Esta
morsa vivió hace 15 millones de ańos, en un momento de
calentamiento global en la Tierra.
Morenocetus
parvus. Cabrera, 1926.
Cráneo de Morenocetus
parvus de la colección del Museo de La Plata.
Prensa.
Aspecto en vida de Morenocetus parvus por el
paleoartista Jorge Gonzalez.
Argentina. Se puede distinguir del
único otro balaenido mioceno, Peripolocetus,
por tener un proceso cigomático expandido. La longitud del
cuerpo de Morenocetus se estima en unos 5
metros, alimentándose de pequeńos invertebrados marinos. El
primer balaenido reconocido es el Mioceno temprano
Morenocetus parvus en Argentina. Se describió
brevemente a partir de dos cráneos incompletos, una
mandíbula y algunas vértebras cervicales recogidas de la
Formación Gaiman del Mioceno inferior de la Patagonia. Desde
entonces no ha sido revisado, por lo que sigue siendo un
cetáceo fósil frecuentemente citado pero enigmático con gran
potencial para arrojar luz sobre la historia temprana de la
corona Mysticeti. Pero estudios recientes, demuestran que Morenocetus es el balaenido extinto más antiguo
hasta ahora, aunque un espécimen chaeomysticete de depósitos
marinos del Oligoceno tardío en Nueva Zelanda como un
posible representante en la línea evolutiva de los
balaenidos. La optimización de la longitud corporal en
nuestra filogenia de Balaenidae sugiere que la condición
primitiva fue una longitud corporal relativamente pequeńa
representada por Morenocetus, y que el
gigantismo se ha adquirido de forma independiente al menos
dos veces (en Balaena mysticetus y
Eubalaena spp.) con la aparición más temprana de
este rasgo en el Plioceno tardío Mioceno-temprano
representado por Eubalaena shinshuensis.
Mandíbula de
Megalodon recreada a tamańo natural en el Museo
Paleontológico de Bariloche.
Diente del
tiburón Carcarocles megalodon, común del Mioceno de
Patagonia y Entre Ríos, Argentina. (*)
Reconstrucción
del gigantesco Carcarocles megalodon.
(*)
América, Asia, Europa, Africa y Oceania.
Durante el periodo Mioceno, en lo que hoy es el Cerro Gualicho en la
Provincia Patagonica de Rió Negro, entre 10 y 12 millones de ańos
atrás, el mar ingreso numerosas veces al continente, depositando
sedimentos con restos fósiles de vertebrados e invertebrados
marinos muy llamativos, sobre todo por sus dimensiones. Carcarocles
megalodon, fue un Selaceo (Tiburón) del cual solo se han preservado
como fósiles sus enormes dientes, ya que al igual que sus
descendientes tenían un esqueleto cartilaginoso, el cual no se
preservo en los sedimentos. Pero los estudios de anatomía comparada
realizados en los últimos ańos brindaron muchos datos importantes.
Carcarocles pudo tener unos 20 metros de largo y pesar hasta 50
toneladas. Los paleontólogos creen que se extinguió hace 2,5
millones de ańos, durante el inicio del Pleistoceno. Otros piensan
que podrían vivir en la actualidad en aguas muy profundas, al igual
que las especies de tiburones descriptas recientemente. Carcarocles
megalodon, tenia unos 45 kilogramos de dientes, los cuales
variaban su tamańo desde unos pocos centímetros hasta los 20
centímetros y cada uno pesaban 300 gramos. Se alimentaban
principalmente de ballenas, lobos marinos primitivos y grandes
peces. Sinonimia: Procarcharodon megalodonCasier, 1960 y Megaselachus megalodon
Glikman, 1964.
Megapiranha paranensis.
Cione et al. 2009.
Fragmento mandibular. (*)
Fragmento mandibular. (*)
Reconstrucción. (*)
Argentina.
En el Museo de La Plata
revelan un enigma paleontológico. El ejemplar medía más de un metro.
Que hace unos 10 millones de ańos los ríos de la Mesopotamia
Argentina, estaban habitados por una megapirańa -un pez carnívoro de
más de un metro de largo- era algo que se suponía desde hace tiempo.
Sin embargo hasta el momento no había pruebas de su existencia. Fue
un investigador platense quien las halló; no en un trabajo de campo,
sino revisando antiguas cajas con fósiles indiferenciados en los
depósitos del Museo de Ciencias Naturales de La Plata. La reciente
publicación de su hallazgo despertó gran interés, tanto en el ámbito
de la paleontología como en el de la divulgación científica.
Revistas como la National Geographic se hicieron eco del
descubrimiento del doctor Alberto Cione, paleontólogo del Museo de
La Plata e investigador del Conicet. Es que su hallazgo vino a
resolver además un misterio paleontológico: el del salto evolutivo
que separa a las pirańas actuales con el pacú, un pariente herbívoro
mucho más primitivo. Aunque se conoce ahora, la historia del
descubrimiento se remonta a fines de los '80, cuando Cione -a cargo
de la colección de peces de la división de Paleontología de
Vertebrados- revisaba antiguas cajas con material fósil sin
identificar. Esas piezas habían sido desenterradas de las barrancas
del Río Paraná, cerca de la localidad de Villa Urquiza, hacia el
1900, y desde entonces permanecían guardadas. Ancestro directo de
las actuales pirańas, aunque de un tamańo cinco veces mayor, las
megapirańas habrían habitado los ríos, y quizás también las lagunas,
de la mesopotamia hace entre 8 y 15 millones de ańos. Nadie está
seguro, sin embargo, de qué se alimentaban exactamente. Es posible
que su dieta fuera diversa, como la de las pirańas actuales, que si
bien son eminentemente carnívoras, también comen plantas y frutos.
Lo que sí se sabe es que coexistían con enormes bagres, delfines de
agua dulce, tortugas y cocodrilos de muy diversos tipos, incluso uno
labial de rostro alargado; toda una fauna que se extendía por una
amplia región geográfica hasta el Amazonas. Pese a que las únicas
pruebas de su existencia se hallaron en nuestro territorio, es muy
probable que las megapirańas hayan llegado a habitar también gran
parte de los ríos de Sudamérica. Por entonces las temperaturas
globales eran mucho mas altas que ahora, y las cuencas del Paraná y
el Amazonas no estaban, como hoy, aisladas una de otra.
Ophiocrossota kollembergorum.
Caviglia, Martínez & del Río, 2007.
Ophiocrossota kollembergorum, hallado en Punta
Maqueda. Formación Chenque, en la provincia de Chubut. (*)
Aspecto de
un Ophiuroidea.
(*)
Argentina.
Los ofiuroideos son una clase de equinodermos).
Presentan simetría pentarradial y tienen un aspecto parecido a las
estrellas de mar (asteroideos), con cinco brazos que salen de un
disco central. Seguramente, ofiuroideos y asteroideos son grupos
hermanos. Un fósil muy interesante corresponde justamente a Ophiocrossota
kollembergorum(Ophiuroidea), Punta Maqueda. Formación
Chenque , en la provincia de Chubut. Tienen el cuerpo pequeńo y
aplanado formado por un disco redondeado por cinco brazos
articulados muy finos y largos que surgen bruscamente del disco
central y pueden estar ramificados. La boca está compuesta por cinco
placas móviles que actúan como mandíbulas. Carecen de ano, por lo
que su tubo digestivo es incompleto. Los materiales no digeridos se
expulsan por la boca. El esqueleto es de gran similitud al de las
estrellas de mar, salvo por la presencia de vértebras esqueléticas
en los brazos, que reducen u obliteran completamente la cavidad
celomática.
América del Norte (Nebraska).
Tenia 80 cm de longitud Así como
Phlaocyon tenía un
parecido superficial con los felinos, Phlaocyon se
asemejaba más a los mapaches (prociónidos). Sin
embargo, por varios rasgos del cráneo resulta evidente que
pertenecía a la familia de los perros (cánidos), si bien
era un miembro muy primitivo.Es probable
que Phlaocyon
leucosteus
viviera de
una forma bastante similar a los mapaches actuales. Si bien
tenía las patas típicas de los perros, los miembros estaban
adaptados para subir a los árboles, más que para correr.
Tenía la cabeza corta y ancha, con los ojos dispuestos en la
parte anterior.La mandíbula
superior de esta criatura era curva, como la del mapache, y
tanto los premolares como los molares estaban hechos para
moler, y no incluían las cuchillas para desgarrar la carne
que caracterizan a los perros. Esta dentadura permite
suponer que
Phlaocyon
era omnívoro y que tenía una dieta combinada
de semillas, frutos, insectos y huevos de aves, además de
mamíferos pequeńos y aves.
América del Norte {costa delPacífico).Tenia 1,5 m de
longitud. Este mamífero marino de aspecto primitivo
representa una de las primeras etapas de la adaptación de un
carnívoro terrestre a una forma de vida marina.
Enaliarctos
mealsi,
se encuentra casi a mitad de
camino entre la nutria y el león marino. Sus muelas todavía
tenían cuchillas para desgarrar la carne (carniceras), como
las de los perros terrestres. El cuerpo era fusiforme y
bastante parecido al de la nutria, con patas definidas y
cola, aunque los pies ya se habían modificado para
convertirse en aletas.
Es probable que Enaliarctostuviera una manera de vivir bastante parecida a la de la
nutria de mar moderna, pasando parte del tiempo sobre la
tierra, además de en el mar, y consumiendo una variedad de
animales marinos, incluidos tanto los peces corno los
crustáceos. Sin embargo, ya habían aparecido algunas de las
características de los leones marinos, como los ojos
grandes, los sentidos complejos asociados con los bigotes y
la especialización del oído interno para detectar la
dirección del sonido debajo del agua. Todos estos sentidos
le servían a Enaliarctospara localizar
a sus presas. Es probable que el olfato tuviera un papel
insignificante para cazar, como ocurre en los pinnipedos
actuales.
Panamá. América Central.
El hallazgo reciente de fósiles muestra que pequeńos
camellos con grandes hocicos deambularon por la selva
tropical de Panamá hace unos 20 millones de ańos. Un
estudiante de doctorado de geología de la Universidad de
Florida encontró los fósiles de camello cuando analizaba el
sedimento de una zona de obras en el Canal de Panamá. La
especie no tiene joroba, a diferencia de los actuales
rumiantes y una de las dos especies encontradas al parecer
se levantaba en dos patas. Los investigadores creen que los
camellos, Aguascalientia panamaensis y
Aguascalientia minuta, posiblemente utilizaron sus
afilados dientes para masticar follaje exuberante y fruta.
El hallazgo está despertando dudas sobre hace cuánto tiempo
se creó el istmo y el descubrimiento de un fósil de mamífero
puede ayudar a los científicos a entender mejor qué sucedió
cuando las Américas del Norte y el Sur finalmente se
conectaron. Aunque es un país relativamente pequeńo, Panamá
tiene gran importancia científica porque sirve como el
puente terrestre que une al continente. Cuando se creó esta
puerta entre los continentes, hubo un cambio global en los
mares: los océanos Pacífico y Atlántico se separaron y
comenzó un gran intercambio de animales, llevando a algunas
especies a la extinción y otras a adaptarse. Los científicos
pensaron durante mucho tiempo que el istmo se creó hace 3,5
millones de ańos, pero ahora han descubierto especies de
camellos que vivieron en el área unos 17 millones de ańos
antes
Europa (Francia), África (Kenia), Asia (Pakistán) y América del Norte
(Nebraska). Tenia 3 m de altura Este mastodonte
de cuatro colmillos estaba muy difundido y sus fósiles han
aparecido en cuatro continentes. Como consecuencia de esto,
el mismo animal fósil ha recibido varios nombres, entre los
que se incluyen Trilophodon y
Tetrabelodon.
La mandíbula inferior, con sus colmillos paralelos, era muy
larga. Probablemente se utilizaba en combinación con una
trompa igual de larga, situada en la mandíbula superior.
Hubo una reducción progresiva de la cantidad de dientes,
pero los que quedaron desarrollaron una cantidad de
rebordes elevados, o cúspides, para aumentar la superficie
donde se molía. Esto era necesario para procesar las enormes
cantidades de alimentos vegetales que se necesitaban para
satisfacer a un animal de semejante corpulencia. La mayoría
de los miembros del género Gomphotherium
comían las hojas de los arbustos, pero una especie vivía en
las ciénagas y se aumentaba de plantas acuáticas tiernas.
América del
Norte (Colorado,Nebraska). Tenia 3 m de
altura.
Amebelodon era un
miembro típico de !a familia de los gonfotéridos,
alto y con colmillos en forma de paletas, que ramoneaba por
las praderas de América del Norte, durante el final del
Mioceno. Hace alrededor de 10 millones de ańos, los bosques
se habían convertido en vastas praderas secas, un ambiente
más adecuado para los animales corredores ungulados, como
los caballos.Pero también atravesaban las planicies
muchos ríos sinuosos en los que abundaban las plantas
acuáticas exuberantes. En este nicho se desarrolló
Amebelodon.Por
el tamańo de su cuerpo y la apariencia general,
Amebelodonse parecía a los elefantes modernos;
aunque el cráneo y los colmillos eran muy diferentes. Los
colmillos aplanados de la mandíbula inferior, alargada,
estaban el uno junto al
otro, formando una especie de pala que se proyectaba más de
1 m y tenía un borde cortante como el de una espada.Se
supone que Amebelodon sujetaba las plantas
acuáticas que le servían de alimento entre la trompa
aplanada y los colmillos, las arrancaba del barro del fondo
del río y después las empujaba con la trompa a lo largo de
la pala, hasta introducirlas en la boca.
Thalassoleon
mexicanus.
Repenning y Tedford, 1977.
México y Estados Unidos.
Las focas, los leones marinos y las morsas
evolucionaron de antepasados similares a osos en el
oligoceno (33,7-23,5 millones de ańos atrás). Los primeros
tipos se parecían a las nutrias, pero tenían manos y pies
con forma de aletas y grandes ojos que les permitían ver
mejor dentro del agua. Para fines del mioceno, las focas se
habían diseminado por el mundo, mientras que las morsas y
los leones marinos, como el Thalassoleon,
evolucionaron en el hemisferio norte.
Espańa, Europa. Este
grupo de mamíferos, que se caracterizan porque las crías
completan su desarrollo en el interior de la bolsa marsupial
materna (o marsupio), habitaron también en épocas pasadas en
África, Asia y Europa. En estos tres continentes, los
marsupiales se fueron extinguiendo a lo largo del Terciario,
y el último registro en Europa data del Mioceno. La especie
fósil hallada en tierras castellonenses, cuyo nombre
científico es Amphiperatherium frequens, está
emparentada con los marsupiales americanos, ya que forma
parte de la
familia de los didélfidos, a la que también
pertenecen las zarigüeyas actuales.
El ambiente donde vivían estos animales debió tratarse de
una zona boscosa subtropical, en las cercanías de un gran
lago
que abarcaba los actuales términos municipales de Alcora y
Ribesalbes. En la parte profunda del lago se depositaron los
sedimentos que constituyen el clásico yacimiento
paleontológico de conservación excepcional de Ribesalbes,
que ha proporcionado "magníficos" fósiles de plantas,
insectos y anfibios. En una etapa posterior, en los fangos
de las riberas del lago se acumularon los restos
esqueléticos de mamíferos que constituyen los yacimientos de
Araia. Para su obtención, se tuvieron que procesar varias
toneladas de sedimento, que fue tamizado para eliminar las
arcillas y los limos. Posteriormente, se procedió a triar el
residuo grano a grano para extraer los fósiles, que
consisten sobre todo en dientes aislados. Para
su estudio se requieren técnicas de microscopía electrónica,
ya que estas piezas dentarias poseen un tamańo que no suele
superar el milímetro.
Murcia, Espańa. Es una
especie de tortuga terrestre gigante, cuyos caparazones
superan el metro de diámetro y pesan más de una tonelada,
que datan de hace más de seis millones de ańos. El hallazgo
de los cuatro caparazones, dos de ellos todavía sin extraer,
se produjo el pasado octubre durante las obras de
construcción del tercer carril de la autovía A-31, que une
el Puerto de la Cadena con el polígono de San Ginés, por
parte de la empresa constructora, y fue certificado por la
Asociación Cultural Paleontológica Murciana. Los trabajos
llevados a cabo por los técnicos han conllevado también la
recuperación de 35 fragmentos de fósiles y huesos de bóvidos
y caballos, entre otros animales, como falanges, vértebras,
dientes o costillas. La hipótesis de que hace más de 11
millones de ańos la zona estaba cubierta por el mar, poco a
poco fue saliendo a la superficie y surgió un delta que fue
introduciendo sedimentos desde el mar que arrastraron los
caparazones de las tortugas muertas al lugar donde se han
encontrado.
Europa (Montańas del
Cáucaso), Asia (Mongolia) y África (Kenia).
Tenía
3 m de altura. Platybelodon
era otroanimal con colmillos en forma de paletas, similar a
Amebelodon,
que vivió en Europa y Asia. Tenía las paletas
mas cortas y mas anchas
estaban hendidas a ambos lados para dejar
lugar a los colmillos de la mandíbula superior. Es evidente
que el estilo de vida de Platybelodonera muy
parecidoal de Amebelodon:
se bańaba en aguas poco profundas y arrancaba
plantas acuáticas.
El hecho de que en esta época vivieran en
Eurasia y América del Norte distintos tipos relacionados de
proboscídeos indica la existencia de una ruta migratoria
entre estas dos masas continentales, que atravesaba un istmo
que había donde hoy está el estrecho de Bering. Los animales
con colmillos en forma de paletas presentan una
espedaliíadón excesiva para un tipo determinado de
alimentación. Como ocurre con las demás criaturas demasiado
especializadas, eran sumamente vulnerables a los cambios
ambientales y no vivieron durante demasiado tiempo.
Simbakubwa
kutokaafrika. Borths
& Stevens, 2019.
África (Kenia).
Es una especie y género extintos de mamífero perteneciente a
la familia Hyainailouridae, comprendida en el orden Hyaenodonta.
Vivió a principios del Mioceno y sus restos fueron hallados
en Kenia. El espécimen tipo fue recuperado de depósitos
datados en 23 millones de ańos de antigüedad. Con un peso
estimado en alrededor de 1,500 kilogramos, puede haber
sobrepasado al actual oso polar en tamańo. Los fósiles de Simbakubwa fueron
descubiertos originalmente por Matthew Borths y Nancy
Stevens cuando estaban examinando los fósiles alojados en
el Museo Nacional de Nairobi en Kenia. Simbakubwa,
como otros hienailoúridos, probablemente era un depredador y
carrońero especializado que se alimentaba de grandes
mamíferos como rinocerontes y proboscídeos primitivos.
Parece haber estado algo menos adaptado en triturar huesos
que parientes suyos posteriores tales como Hyainailouros.
Sin embargo, al igual que este último, Simbakubwa poseía
bordes cortantes girados en sentido lingual en sus dientes carnasiales,
lo que aseguraba que el borde cortante se mantuviera
constante a través de su vida.
Anoia, Catalunya, Europa. Paleontólogos del
Institut Catalŕ de Paleontologia (ICP)
han descubierto una nueva especie de castor que vivió en Hostalets
de Pierola (Anoia, Catalunya) hace doce millones de ańos.
La nueva especie descrita, Chalicomys batalleri, ya tiene
el fémur como el del
castor actual, que le permite impulsarse usando las patas
posteriores y así se puede mover con rapidez en el agua. Los
castores son los roedores más bien adaptados a vivir en el agua,
hasta el extremo que, mediante la construcción de presas con
troncos, llegan a inundar extensas áreas de bosque para no tener que
alejarse para alimentarse. No obstante, los primeros castores, que
vivieron hace unos 30 millones de ańos, eran animales terrestres que
excavaban madrigueras, y no fue hasta hace unos 25 millones de ańos
que aparecieron algunas formas adaptadas a vivir en el agua pero que
nadaban de una manera mucho menos eficiente que las actuales, usando
las cuatro patas a la vez. La especie se ha
denominado Chalicomysbatallerien
memoria del padre Ramón Bataller, uno de los pioneros de la
paleontología en Catalunya que, además, fue uno de
los primeros especialistas que estudió los fósiles encontrados en
los
Hostalets
de Pierola ya en la década de 1930.
África
(Namibia), Asia (China) y Europa (Francia).
Tenia
2 m
de longitud Si bien ya no quedan osos en África, sí los hubo
en el pasado. Agriotherium
vivió en el sudoeste de África, que entonces
era un continente completo, separado de Europa y de Asia.
Agriotheriumera un oso
muy grande, mayor incluso que el oso gigante de Kodiak.
También era muy primitivo y se parecía bastante a los
perros, en algunos sentidos. Sin embargo, los dientes se
ajustaban al modelo típico de los osos.Por
consiguiente, parece seguro suponer que se trataba
de un omnívoro.
África. Asia y Europa.
Tenia 1,5 metros de longitud. Las hienas del género
Percrocuta fueron las más grandes que existieron.
Una de las especies, Percrocuta gigantea,
oriunda de China, era tan grande como el león actual. A
pesar de su enorme tamańo, Percrocuta era muy
similar a la hiena manchada, Crocuta crocuta,
que vive actualmente en África. Esta criatura estaba mucho
más difundida durante el Pleistoceno que en la actualidad;
se han encontrado fósiles por toda África, Europa y Asia.
Como su familiar moderno, Percrocuta tenía una
cabeza grande y unas mandíbulas muy poderosas, armadas con
unos dientes enormes, capaces de triturar huesos. Adoptaba
la típica postura inclinada de sus familiares actuales, con
las patas delanteras más largas que las traseras.
África (Marruecos), Europa (Grecia)
y Asia. Tenia 1,2 metros de longitud. Aunque las
hienas modernas puedan parecerse más a los perros que a los
gatos, pertenecen al mismo grupo de carnívoros que éstos.
Las características esqueléticas muestran que las hienas
primitivas eran similares a las civetas y jinetas. Estas
hienas moledoras de huesos evolucionaron en el mioceno y se
diseminaron por África, Asia y Europa. Las hienas
primitivas, como el Ictitherium, eran mucho
más pequeńas que las modernas. Seguramente fue un gran
depredador y un carrońero oportunista muy tenaz.
Ictitherium fue una de las hienas más primitivas y
es probable que se pareciera más a la civeta, por su
estructura y su aspecto. Sus dientes también eran como los
de la civeta, adecuados para una alimentación a base de
insectos, en lugar de los formidables dientes de las hienas,
que les servían para destrozar huesos. Junto con sus
familiares primitivos, Ictitherium fue uno de
los cazadores más difundidos de la época. Incluso, durante
una etapa del Plioceno, sus restos fósiles superan en número
a los de todos los demás carnívoros en conjunto. A menudo se
encuentran grupos de animales que se han fosilizado juntos,
lo que indica que una inundación los ha arrastrado al mismo
tiempo. Es probable que esta hiena primitiva ya hubiese
desarrollado un orden social bastante avanzado y que cazase
en manadas, como hacen las hienas actuales.
Torrejón de Velasco, Espańa.
Fueron depredadores dominantes hace 9 millones de
ańos, pero su propia especialización los llevó a la
extinción. Fósiles hallados en el Cerro de Batallones ayudan
a explicar la evolución de los «tigres con dientes de sable
de europa» o macairodontinos.
Son criaturas extinguidas
desarrollaron un método de caza más rápido y seguro, con
ventajas sobre el de los felinos que matan a sus presas por
asfixia. Y es que, como afirma Salesa, «los grandes dientes
y otros rasgos de los primeros macairodontinos les
permitieron cazar empleando menos tiempo y energía, y con
menos riesgo de sufrir heridas o fracturas durante la
inmovilización de la presa». Dada la escasez de restos
fósiles de félidos macairodontinos, el yacimiento de
Batallones es un filón que poco a poco va ofreciendo a la
comunidad científica pistas y datos sobre el origen
evolutivo de una especie que, a pesar de su nombre común, no
es antepasada de ningún felino. Para Salesa, investigador de
la anatomía, filogenia, biomecánica y paleoecología de
fósiles del Neógeno, especialmente félidos
macairodontinos, el estudio de los 24 fósiles de
Paramachairodus ogygia -uno de los
macairodontinos más primitivos y del tamańo de un puma-
hallados en Batallones desvela lo que hasta ahora suponía
uno de los grandes misterios de la especie: cuál era su
anatomía, cómo se movían y cómo mataban a sus presas. Así,
el análisis de los fósiles revela el inicio del desarrollo
de grandes caninos aplanados, unido a una serie de
modificaciones craneales, mandibulares y cervicales. Por
ello, los Paramachairodus ogygia inmovilizaban
a sus presas y con una fuerte flexión de la cabeza les
clavaban los caninos de sable en la garganta, lo que suponía
el desmayo y una muerte rápida al cortar el suministro de
sangre al cerebro. Como reconocen los investigadores del
Museo Nacional de Ciencias Naturales de Espańa, esta táctica
les convirtió rápidamente en cazadores dominantes que
siguieron evolucionando para poder apresar animales del
tamańo de cebras, jirafas y ńúes. Pero esa misma
especialización pudo ser la causa de su extinción. En
palabras de Salesa, «una de las posibles causas fue un
cambio en las faunas de herbívoros, que hizo que durante
cierta época del ańo las presas migraran, como hacen los
ńúes en África. Los macairodontinos tendrían
problemas para alimentarse de animales más pequeńos y así
fueron haciéndose más escasos y desaparecieron. El último de
los «tigres con dientes de sable» fue el Smilodon,
que vivió en América hasta hace 10.000 ańos. Sin embargo,
los fósiles de Batallones eran sus antepasados, por lo que
este hallazgo abre nuevas vías de investigación y podría
revelar nuevas claves sobre los orígenes de otros grandes
depredadores.
Elche,
Espańa. Los restos del animal, que posee cuatro colmillos de
dos metros, fueron hallados cerca de Elche y podrían
desvelar una nueva especie. Paleontólogos del Museo
Paleontológico de Elche (MUPE) excavaron entre julio y
octubre de 2006 en el sur de Alicante los restos de un
elefante prehistórico completo de 8,5 millones de ańos que,
con cuatro colmillos, podría pertenecer a una especie nueva.
El coordinador del equipo de excavaciones, el paleontólogo
Enrique Peńalver, informó de que podría tratarse de un
ejemplar de Proboscidio del género hallado en un yacimiento
del sur de la provincia de Alicante. Se trata de un animal
de tamańo similar a un elefante africano actual pero con
cuatro colmillos o defensas, de unos dos metros de longitud
cada una de ellas. Este ejemplar parece ser que vivió
durante el periodo Mioceno, concretamente en la fase
Vallesiense de esta época glaciar. En el yacimiento se
encuentra el animal completo aunque algo disperso, según
Peńalver. En esta línea, el coordinador indicó que estos
animales llegaron a la actual Península Ibérica procedentes
del continente africano “a través del estrecho de Gibraltar
que, por aquel entonces, no estaba cubierto por el mar”, a
lo que ańadió: “invadieron Espańa y Europa, así como la
provincia de Alicante, que era un área muy cálida pero que
tenía también cursos de agua, por lo que presentaba un
paisaje similar a las actuales partes áridas de Africa”. Por
su parte, el director del MUPE, José Manuel Marín, afirmó
que este importante hallazgo servirá para ampliar las
investigaciones y la información que se tiene actualmente
sobre ese periodo geológico. En ese mismo yacimiento ya se
hallaron en el pasado restos de animales de la misma época,
como tigres de dientes de sable, tortugas de gran tamańo y
puercoespines prehistóricos, a los que se une ahora los
restos de este gran elefante.
Europa
(Alemania y Bohemia), Asia (India) y África (Kenya).
Tenia 4 m de altura. La característica más destacada de este
elefante eran sus colmillos, aunque todavía se sigue
discutiendo su utilidad. No había ningún colmillo en la
mandíbula superior, pero la inferior estaba doblada hacia
abajo, en ángulo recto, y de allí surgían dos inmensos
colmillos curvos. Esta forma parecía tan poco probable que,
cuando en la década de 1820, los científicos trataron de
realizar las primeras reconstrucciones de Deinotherium,
le pusieron la mandíbula al revés. Es posible que el
animal utilizara los colmillos para arrancar la corteza de
los árboles o para desenterrar tubérculos. Si logró
sobrevivir casi sin cambiar durante 20 millones de ańos es
evidente que Deinotherium era un animal muy próspero.
Europa (Bélgica y
URSS) y Argentina.
Tenía 4 metros de longitud. Cetotherium pertenecía a una familia de ballenas
primitivas que evolucionó a fines del Oligoceno y alcanzó su
auge durante el Mioceno, hace unos 15 millones de ańos.
Tenía un parecido sorprendente con la ballena gris actual
del Pacífico Norte, aunque tenía menos de la tercera parte
de su longitud.Es probable que sus placas
fuesen bastante cortas, aunque esto resulta difícil de
determinar, ya que las ballenas, como los cuernos y los
pelos, no se fosilizan. Sin embargo, los cráneos conservan
las marcas de los vasos sanguíneos que aumentaban a las
ballenas durante su vida, y a partir de estos restos se
pueden averiguar muchas cosas.Cetotherium
riabinini y sus
familiares eran, probablemente, presas de una especie de
enorme tiburón blanco, Carcharodon,
que, a juzgar por el tamańo de los dientes que a menudo se
encuentran fosilizados, alcanzaba un tamańo similar al de
una ballena pequeńa.
América del Norte,
Europa y
Asia.Al
principio de su historia, los cerdos eran mucho más variados
y estaban más extendidos que ahora. Uno de los primeros
grupos, de los bosques de América del Norte, Europa y el
oriente de Asia de hace 25 a 50 millones de ańos, fue el de
los entelodontes, como el Archaeotherium
mortoni. El
cráneo era largo y tenía dientes afilados al frente, entre
ellos dos colmillos y muchas muelas anchas detrás. Las
mandíbulas eran profundas, lo que muestra que debían tener
músculos fuertes. También tenía grandes protuberancias óseas
en la mandíbula inferior cuya función se ignora. El
Archaeotheriumprobablemente se alimentaba de
plantas duras y raíces. La zona del cerebro relacionada con
el sentido del olfato estaba muy desarrollada, por lo que el
Archaeotheriumtal vez era capaz de oler
raíces desde la superficie. Tenía cuello corto por el gran
tamańo de su cabeza: cabezas grandes requieren grandes
músculos para moverlas . Las patas eran lo suficientemente
grandes como para correr entre los árboles.Los entelodontes se extinguieron cuando
las praderas se extendieron en América del Norte, Europa y
Asia, pues probablemente no estaban adaptados para vivir en
terreno abierto.
Asia (Mongolia), Europa (Francia y
Espańa) y América del Norte (Estados Unidos). No
obstante su enorme tamańo, Hemicyon era
bastante ligero para ser un oso. En realidad, se parecía más
a un perro pesado; su nombre quiere decir «medio perro».
Probablemente, era más carnívoro que la mayoría de los otros
osos y se supone que era un cazador activo. Tenía piernas
fuertes y la estructura de los pies indica que corría sobre
los dedos (una adaptación para correr a toda velocidad) en
lugar de hacerlo apoyando todo el pie en el suelo, como los
osos actuales. De estas características se deduce que
Hemicyon era un cazador de vastas planicies y es
posible que vagara en grupos.
Megantereon
nihowanensis. Teilhard
de Chardin & Piveteau, 1930.
África (Sudáfrica), Asia (India), Europa (Francia) y América
del Norte (Texas). Tenia 1,2 metros de longitud.
Megantereon era un
auténtico felino de " dientes de sable primitivo, y es
probable que fuese el antepasado de otras formas. Los
dientes no eran lo bastante largos como para parecer
verdaderos sables; más bien parecían dagas, por su tamańo y
su forma, y a esto se debe el que Megantereon
y sus familiares inmediatos a menudo se conozcan con el
nombre de felinos de dientes de daga. Gracias al desarrollo
de largos caninos, estos predadores poderosos podían matar a
los mamíferos de piel gruesa que pacían en el mismo hábitat
que ellos.
Megantereon prosperó en la
región mediterránea durante el final del Plioceno y el
principio del Pleistoceno, entre tres y dos millones de ańos
atrás. También se difundió por África y América de! Norte,
desde su origen, en el norte de India, a fines Mioceno.
Asia (Japón) y América del Norte
(California y Oregón). Tenia 1.7 m de longitud.
La típica forma fusiforme del león marino actual había
comenzado a aparecer con Desmatophoca. Igual
que en sus familiares vivos, los miembros anteriores eran
más poderosos que los posteriores, y los pies estaban
modificados para formar aletas, con los dedos alargados,
desplegados y unidos mediante unas membranas de piel, para
producir una superficie amplia para nadar. Todos los huesos
de las extremidades se habían acortado para darles más
fuerza. Aunque Desmatophoca brachycephala todavía tenía
cola, en contraste con los leones marinos, ésta se
encontraba muy reducida y no era más larga que el cráneo del
animal. Como su antepasado Enaliarctos, tenía
unos ojos enormes, lo cual indica que la vista era el
sentido más importante para cazar. Es posible que no tuviera
el oído totalmente adaptado para percibir los sonidos
submarinos, pero no hay duda de que le servía cuando se
encontraba en tierra.
Asia y América del Norte (las
costas del Pacífico).Eurhinodelphis
tenia 2 metros de longitud. Los odontocetos se dividieron en
varios grupos durante el Oligoceno, hace unos 30 millones de
ańos.
Eurhinodelphis
era un miembro típico de la familia de los rabdosteidos, que
incluye a las marsopas de hocico largo. La estructura de los
oídos se había vuelto mucho más compleja que la de los
odontocetos más primitivos, de modo que es probable que esta
ballena hubiese desarrollado el complejo sistema de
detección a través de ultrasonidos que se observa en las
ballenas dentadas actuales. Las ballenas vivas utilizan una
forma de sonar ultrasónico; emiten sonidos ligeros de alta
frecuencia que rebotan en los objetos y los ecos se analizan
en su enorme cerebro para determinar, con sorprendente
precisión, el tamańo, la forma, la distancia y la velocidad
de un objeto y, por supuesto, si es comestible o no. Al
igual que el del delfín moderno, el cráneo de
Eurhinodelphis se
ha vuelto un poco asimétrico y las estructuras que aparecen
en uno de los lados son diferentes a las que aparecen en el
otro. Esta distribución se podría vincular al desarrollo de
nuevas capacidades, como la de perseguir presas de
movimientos rápidos y navegar con mayor exactitud. Sin
embargo, la característica mas peculiar de
Eurhinodelphis
era su hocico alargado. No tenía dientes en el extremo y el
posible que empleara la «espada» del pez espada actual para
golpear y aturdir a sus presas, que entonces quedaban
atrapadas en las mandíbulas del animal.
Bullockornis planei.Rich,
1979.
Australia, Oceanía.
Es un género extinto de ave no voladora de la familia Dromornithidae que vivió a mediados del Mioceno, hace
aproximadamente 15 millones de ańos, en lo que ahora es
Australia. Bullockornismedía aproximadamente
2,6 metros de altura, y puede haber llegado a pesar
500 kilogramos. Las características del cráneo de
Bullockornis, incluyendo un enorme pico adaptado a
cortar, indican que el ave puede haber sido carnívora. Dicho
cráneo es mayor que el de muchos caballos pequeńos. Varios
paleontólogos, incluyendo a Peter Murray del Museo Central
Australiano, creen que Bullockornis estaba
relacionado con los gansos y patos. Esto, unido al enorme
tamańo del ave y a sus posibles hábitos carnívoros, han
originado su notorio apodo. Por su parte, el nombre
científico del género significa "ave buey".
Heracles
inexpectatus. Digno, Mano,
Arquero, Scofield y De Pietri, 2019.
Nueva Zelanda.
Un equipo de paleontólogos australianos ha descubierto los
restos fósiles del loro más grande del mundo hallado hasta
ahora. Se calcula que este singular animal tenía una altura
de un metro de altura y pesaba 7 kilos. Los restos del
ejemplar de la nueva especie han sido encontrados en el
centro de Otago, Nueva Zelanda, concretamente en una zona
muy conocida por albergar diferentes tipos de aves de gran
tamańo. Los expertos lo han llamado Heracles inexpectatus,
nombre que surge de su gran tamańo y fuerza hercúlea y del
carácter inesperado del hallazgo. Los restos tienen una
antigüedad de unos 19 millones de ańos. Sus dimensiones son
el doble de grandes que las del Kakapo neozelandés (Strigops
habroptilus), que es el loro más grande que se conocía hasta
ahora y que está en peligro crítico de extinción. Según los
expertos, el Heracles inexpectatusvivía en un bosque
subtropical donde había muchas especies de laureles y
palmeras.
Nimbacinus dicksoni.
Muirhead & Archer, 1990.
Australia, Oceanía. La especie
Nimbacinus dicksoni forma parte de una familia extinta de
carnívoros marsupiales australianos y de Nueva Guinea, la
Thylacinidae. Si exceptuamos a una especie extinguida en
tiempos modernos, la mayor parte de la información conocida
sobre las especies de esta familia procede de fragmentos de
cráneos recuperados, lo que limita el análisis de su
diversidad y su ecología. Gracias al hallazgo de un cráneo
conservado de N. dicksoni de aproximadamente 16 a 11,6
millones de ańos de antigüedad, procedente del yacimiento
paleontológico de Riversleigh, declarado Patrimonio de la
Humanidad, y ubicado en el noroeste de Queensland, en
Australia, unos especialistas de la Universidad de Nueva
Inglaterra y la Universidad de Nueva Gales del Sur, ambas en
Australia, han resuelto finalmente el enigma de si el N. dicksoni
era capaz de cazar presas grandes o solo pequeńas.
Las similitudes en cuanto a rendimiento mecánico del cráneo
entre el N. dicksoni y el Dasyurus maculatus (quol de cola
moteada o gato tigre) son mayores que con respecto al tigre
de Tasmania. Attard y sus colaboradores sugieren que el N. dicksoni, un marsupial de tamańo medio y un peso de unos 5
kilogramos, poseía una fuerza de mordedura superior para el
tamańo que tenía, era sobre todo carnívoro, y probablemente
era capaz de cazar vertebrados que excedían su propia masa
corporal. El
hallazgo sugiere que el Nimbacinus dicksoni era un cazador
oportunista, cuyas presas potenciales incluían a pájaros,
ranas, lagartos y serpientes, así como un amplio surtido de
marsupiales.
Barnes, Lawrence G. (1988-03-18). "A New Fossil Pinniped (Mammalia:
Otariidae) from the Middle Miocene Sharktooth Hill Bonebed,
California". Contributions in Science (Kansas: Natural
History Museum of Los Angeles County) (396): 1–11.
Berta, A. 1991. New Enaliarctos* (Pinnipedimorpha) from the
Miocene of Oregon and the role of "Enaliarctids" in Pinniped
Phylogeny. Smithsonian Contributions to Paleobiology 69.
Lambert, W. D., and J. Shoshani, 1998. The Proboscidea. In
Janis, C., K. M. Scott, and L. Jacobs (eds.), Evolution of
Tertiary Mammals of North America, Volume 1. Cambridge
University Press: Cambridge, UK.
Magnussen Saffer,
Mariano (2013). Algunos
representantes característicos de la fauna del Mioceno. Paleo,
Revista Argentina de Paleontología. Boletín Paleontológico.
Ańo 11. 95: 25-31.
Magnussen
Saffer, Mariano (2010).
Algunos integrantes de la paleofauna del Terciario del
territorio argentino.
Paleo,
Boletín Paleontológico. Ańo 8. 47: 22-23.
Magnussen Saffer,
Mariano. 2015. Los
mamíferos dientes de sable y su paso en la historia
biológica. Paleo Revista Argentina de
Paleontología. Boletín Paleontológico. Ańo XIII. 132: 13-18.
Magnussen Saffer,
Mariano (2013). Vertebrados del
Mioceno en distintos continentes. Paleo, Revista Argentina de Paleontología. Boletín
Paleontológico. Ańo 11. 96: 32-40.- Magnussen Saffer,
Mariano (2013). Algunos
representantes característicos de la fauna del Mioceno. Paleo, Revista Argentina de Paleontología. Boletín
Paleontológico. Ańo 11. 95: 25-31.
Palmer, D., ed (1999). The Marshall Illustrated Encyclopedia
of Dinosaurs and Prehistoric Animals. London: Marshall
Editions. p. 312.
R. L. Carroll. 1988. Vertebrate Paleontology and Evolution.
W. H. Freeman and Company, New York 1-698R. L. Carroll.
1988. Vertebrate Paleontology and Evolution. W. H. Freeman
and Company, New York 1-698.
Rich, P. (1979) The Dromornithidae, an
extinct family of large ground birds endemic to Australia.
Bureau of National Resources, Geology and Geophysics
Bulletin 184: 1–196.
Rodolfo Salas, José Sánchez, César Chacaltana.
2006. A New Pre-Deseadan Pyrothere (Mammalia) From Northern
Peru And The Wear Facets Of Molariform Teeth Of Pyrotheria.
Journal Of Vertebrate Paleontology, 26(3):760-769
Rodrigues,
J.B. 1892. "Les reptiles fossils de la Vallée de L’Amazone".
Vellosia, Contribuiçőes do Museu Botânico do Amazonas
2:41-60.
W. D. Lambert y J. Shoshani.
1998. Proboscidea. In C. M. Janis, K. M. Scott, and L. L.
Jacobs (eds.), Evolution of Tertiary mammals of North
America 606-621
Ver mas
bibliografía utilizada para hacer la presente Pagina;
AQUÍ.
Ver bibliografía de divulgación sugerida; AQUÍ.
Video;
Best
Documentary of All Time National Geographic
Documentary - Prehistoric Predators: Killer Pig







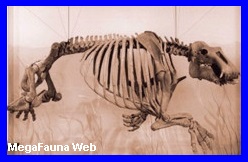















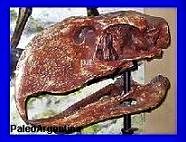



















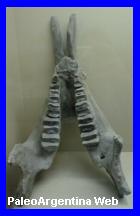
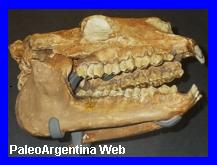




















































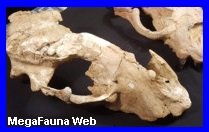






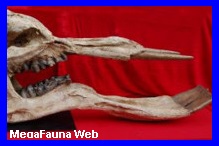

.jpg)
.jpg)