|
Fósiles del Mioceno de
Argentina:
Comprende de siete edades, las cuales comprenden un lapso de 24 a 5
millones de ańos atrás.

Sedimentos de las Barrancas del Rió Paraná.
|
 |
La edad
mamífero "Colhuehuapense", es la más antigua de este
periodo y se encuentra principalmente en la Provincia de
Chubut, Argentina. Posee una antigüedad de 21 a 19 millones
de ańos atrás. La edad "Santacrusense" corresponde a
sedimentos que se han depositado entre 17,5 a 16,5 millones
de ańos, encontrando una amplia distribución en todas las
provincias patagonicas. La edad "Colloncurense" se
caracteriza por su amplio registro fosilífero de vertebrados
de ambientes cálidos y tropicales, marcando drásticamente el
reemplazo de mamíferos sudamericanos. Su antigüedad es de
15,5 a 14 millones de ańos. La edad "Lavarense y
Mayonense" poseen una antigüedad entre 13,8 a 10
millones de ańos. Esta última se encuentra muy bien
representada en las barrancas del Río Mayo, al sur de la
Provincia de Chubut. La edad "Chasiquense" es tal
vez, la más popular del Periodo Mioceno, ya que los
vertebrados extraídos son numerosos. Posee una antigüedad de
10 a 9 millones de ańos y corresponden a los yacimientos
expuestos en las barrancas del Arroyo Chasico (Partido de
Villarino y Adolfo Alsina), en la Provincia de Buenos Aires.
Por último, la edad "Huayquereriense" y corresponden
a los sedimentos depositados entre 9 y 7 millones de ańos
atrás, las cuales afloran a orillas del Rió Paraná y otros
cauces de agua de la mesopotania Argentina y en el NOA, como
Catamarca y Santiago del Estero. En el Mioceno temprano el
clima era muy cálido. La presencia de vertebrados corredores
y pastadores indicarían un ambiente de praderas, con
precipitaciones relativamente abundantes, que alternaban con
ambientes de bosques cálidos. |
|
Para este momento se
produjo una importante emersión en Sudamérica, como
consecuencia de este fenómeno, el mar epicontinental que
bańaba la costa occidental sudamericana registro una leve
regresión. Este fenómeno posibilito en la región del caribe,
la dispersión de algunos pequeńos carnívoros placentarios,
los cuales se encontraban ausentes en América del sur. En la
Antártida las abundantes precipitaciones favorecen la
formación de la capa de hielo. Como consecuencia del periodo
más intenso de orogenia andina comenzó paulatinamente el
crecimiento de la desertización en gran parte del territorio
argentino, sobre todo desde la pampa hasta tierra del fuego.
La elevación de la cordillera de los Andes produjo una
"sombra de lluvias" a lo largo de su flanco oriental,
bloqueando el paso de humedad que acarrean los vientos del
pacifico. Al sur de los 40ş S. Las condiciones al este de la
cordillera pasan de ser bosques cálidos y húmedos a ser
sabanas áridas y templadas por lo que se hacen abundantes
los mamíferos pastadores y ramoneadores. El clima templado y
seco de la Patagonia, favoreció un recambio de los tipos
fáunisticos, que procedió por mucho tiempo a la entrada de
nuevos grupos de mamíferos de origen norteamericano. En la
plataforma continental se desarrolló un mar de aguas
cálidas, someras y cercanas a la costa. A fines del Mioceno,
los cambios climáticos causados por el levantamiento de los
Andes se vieron acentuados por el desarrollo del escudo de
hielo Antártico, lo que produjo un descenso en las
temperaturas costeras y dio origen a corrientes frías que se
hicieron efectivas en la plataforma argentina alrededor de
los 10 millones de ańos atrás. Consecuentemente con esto, la
fauna de las costas argentinas experimento una migración
progresiva hacia el norte, buscado regiones más óptimas para
sobrevivir.<<<
Principales sitios
fosilíferos de Argentina. |
|
Phorusrhacos
longissimus. Ameghino,
1887.
|
|
Phorusrhacos
longissimus
Phorusrhacos
longissimus
Phorusrhacos
longissimus
Phorusrhacos
longissimus
|
|

|
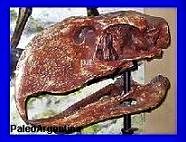 |
 |
|
Cráneo en
el
Museo Jacobacci,
San Antonio Oeste, Rió Negro aportado por la Fundación Azara.
|
Cráneo exhibido en el Museo
Argentino de Ciencias Naturales de Buenos Aires |
Reconstrucción en vivo. Por MarMag.2026 |
Ave.
Durante mucho tiempo, los animales de dieta
carnívora más exitosos en América del sur fueron aves gigantescas,
como los Phorusrhacos. Es un género de gigantescas aves depredadoras
no voladoras que habitaron la Patagonia, de la familia de las
Phorusrhacidae, conteniendo una sola especie; el Phorusrhacos
longissimus. Sus parientes más cercanos en la actualidad son
los de la familia Cariamidae, llamados también Seriemas o Chuńas,
pero su tamańo era mucho mayor que el de éstos; siendo su aspecto
más parecido al de un avestruz. Estas aves habitaron en bosques y
pastizales. Entre los huesos hallados en el estrato de la Formación
Santa Cruz (ahora considerado como principalmente de mediados del
Mioceno) fue la pieza de una mandíbula que Florentino Ameghino
descubierto a principios de 1887 y el mismo ańo la presento como
parte de la cadera de un perezoso por su semejanza, él nombró Phorusrhacos
longissimus. El nombre específico significa "muy largo" en
latín. Lejos estaba Florentino de imaginar aves gigantes en la
antigua Patagonia. El holotipo del ejemplar es, MLP-118 (Museo de La
Plata). En 1889 Ameghino corrigió el nombre a una más correcta
gramaticalmente Phororhacos pero el nombre más
temprana tiene prioridad. En 1891, fue reconocido por él como un
pájaro gigante. Los restos se conocen de varias localidades de la
provincia de Santa Cruz, Argentina. Phorusrhacos situó en torno a
2,5 metros de alto y pesaba unos 130 kilos. Fue apodado el "Ave del
Terror" por razones obvias: es uno de las mayores aves depredadoras
que jamás haya existido, y sus alas rudimentarias formadas como
estructuras de brazo con garras en forma de un gancho de carnicero
para atrapar la presa de frente, y asesinarlo luego con el pico
enorme. El ave mataba de dos maneras. En primer lugar, sujetando a
su presa con su pico y golpear contra el suelo varias veces al igual
que sus parientes modernos, las chuńas. En segundo lugar, y más
espectacularmente, al golpear hacia abajo con la punta aguda en la
parte delantera de su pico superior. En la posición correcta, según
lo especulado, la espiga podría penetrar en el cráneo y matar
instantáneamente. Esta tenía un cráneo enorme, de hasta sesenta
centímetros de largo, armado con un poderoso gancho en la punta del
pico. La estructura del pico y las garras grandes en los dedos
demostrar que se trataba de un carnívoro muy activo. En la Reserva
Natural de Punta Bermejo, en Rio Negro, en sedimentos Miocenos, se
han encontrado una serie de grandes huellas, atribuidas a aves del
terror, asociadas a mamíferos contemporáneos
Kelenken guillermoi. Bertelli et al., 2007.
|

|
 |
 |
|
Esqueleto de
Kelenken recreado por la Fundación Azara a partir de
los restos conocidos. |
Cráneo de
Kelenken guillermoi,
descubierto en Comallo, Provincia de Rió Negro, mide 70
centímetros de largo.
(*) |
Posible
aspecto de
Kelenken guillermoi.
Reconstrucción en vivo. Por MarMag.2026 |
Ave.
Es una especie extinta de ave gruiforme de la
familia Phorusrhacidae o aves del terror. Es el ave predadora
conocida más grande de la historia. Vivió en el Mioceno, hace unos
15 millones de ańos. Midió más de tres metros de altura, su cabeza
(incluyendo el pico) llegó a medir más de 70 cm de longitud, siendo
el ave conocida con el cráneo más grande. Llegaban a pesar los 160
kilogramos. Kelenken era ágil, podía cazar y devorar
roedores, reptiles y pequeńos mamíferos, las crías de estas aves
alcanzaban los 100 km/h pero los adultos sólo llegaban a los 60 km/h.
Su fósil fue encontrado en la Patagonia, concretamente en la
estación de tren de Comallo, cerca de la ciudad de Bariloche, por
Guillermo Oscar Aguirrezabala. Los restos constan de un cráneo,
fragmentos de un pie, de una pata y se encuentran en el Museo
Paleontológico de Bariloche. Antes del descubrimiento de su cráneo,
los científicos interpretaban las cabezas de las grandes aves del
Cenozoico como versiones a mayor escala de las de las aves de menor
tamańo. Los huesos de la pata y el pie tienen un parecido
superficial con los del ńandú, un ave no voladora actual de América
del Sur.
|
Andalgalornis férox.
Patterson
& Kraglievich, 1960. |
|
Andalgalomis férox Andalgalomis férox Andalgalomis férox
Andalgalomis férox Andalgalomis férox Andalgalomis férox |
|

|
 |

|
|
Ejemplar exhibido en el Museo
Argentino de Cs Nat. de Buenos Aires. |
Aspecto del esqueleto de un Ave
del Terror. (*) |
Aspecto de Andalgalornis. Por
MarMag.2026 |
Ave.
Es conocido a partir de un esqueleto incompleto y
algunos huesos aislados encontrados en estratos del Mioceno Superior
al Plioceno Inferior en la Formación Andalgalá, situados en las
provincias de Entre Ríos y Catamarca, en el noreste y norte de
Argentina. Esta ave midió un metro y medio de altura. La subfamilia
a la que pertenecía este género, los Patagornithinae, contenía
especies que eran de constitución más bien delgada; Andalgalornis se
parecía al mucho mayor Phorusrhacos, pero era más
grácil que este, y tenía un pico proporcionalmente más alto. Ademas,
parecia más masivo en proporción a su tamańo corporal entre todos
los forusrácidos. Un análisis realizado con escáner TAC del cráneo
de Andalgalornis sugiere que el pico era fuerte en
sentido dorsoventral (es decir, en el plano vertical) pero en cambio
era relativamente débil mediolateralmente (a los lados). La
debilidad del pico implicaría que esta ave no pudo haber abatido a
presas grandes, sino que depredaba presas más pequeńas, más fáciles
de manejar y con menor riesgo. Aun así, el que, el pico fuera fuerte
dorsoventralmente le pudo haber permitido a Andalgalornis abatir
con rapidez a una presa con una estrategia de caza consistente en
atacar y retirarse de manera repetida. Llego a medir 1,8
metros. Sinónimos: Phororhacos steulleti, Phororhacos
deautieri, Andalgalornis ferox y Phorohacos deautieri.
Rionegrina
pozosaladensis.
Melchor, R. N, et al, 2023.
|
 |
|
 |
|
Rastros o huellas de la icnoespecie de ave del terror
de la Patagonia,
Rionegrina pozosaladensis. |
|
Aspecto de
Rionegrina pozosaladensis.
Por Mar Mag.2026. |
Ave.
En las actuales Áreas Naturales Protegidas de la costa atlántica de
Río Negro —entre Punta
Bermeja, Caleta de los Loros y Pozo Salado— un grupo de guardas
ambientales realizaba sus recorridas habituales, sin imaginar que
estaban a punto de encontrarse con un vestigio único en el mundo.
Durante el
Mioceno tardío, hace
unos 8 millones de ańos, esa región no era como la vemos hoy: era un
ambiente desértico salpicado de extensos lagos que funcionaban como
verdaderos oasis. Allí acudía la fauna de la época a beber, a cazar
o simplemente a sobrevivir. Y fue precisamente en el barro de uno de
esos antiguos lagos donde el pasado quedó detenido en el tiempo. Las
huellas medían casi 40 centímetros de largo. Eran impresiones de un
animal bípedo, que caminaba apoyando principalmente dos dedos: uno
central y otro lateral. Esa característica, poco común, encendió
rápidamente el interés de los especialistas. El productor de esas
pisadas no se parecía a ninguna ave conocida en el registro fósil
mundial. Su modo de apoyo lo diferenciaba de los ńandúes —que
utilizan tres dedos— y también de las chuńas actuales, demasiado
pequeńas para coincidir con el tamańo estimado. A partir del
análisis comparativo, los científicos propusieron que se trataba de
una forma de ave del terror hasta entonces desconocida, a la que
denominaron
Rionegrina pozosaladensis.
Se estima que este animal pesaba al menos 55 kilogramos. Era, sin
duda, un ave corredora, adaptada a la velocidad en terrenos
abiertos. Sus patas estaban diseńadas para desplazarse con eficacia:
un dedo central robusto, otro lateral de apoyo y un dedo interno
reducido que apenas tocaba el suelo. Pero había un detalle aún más
revelador: una gran garra, elevada, que sólo rozaba la superficie en
su punta. Las huellas cuentan más que un simple paso: narran una
escena. El animal caminaba con relativa lentitud, sin huir ni
correr, dejando impresas sus pisadas junto a las de otras especies:
pequeńas aves similares a chorlos, perezosos terrestres y antiguos
parientes del ńandú patagónico. Era un momento cotidiano en un mundo
desaparecido.
Argentavis magnificens. Campbell
& Tonni, 1980.
|
|
|
|
|

|
 |

|
|
Cráneo de Argentavis
magnificens. Foto gentileza de Paleogenesis. |
El Paleontólogo
Rosendo Pascual, comparando el humero de Argentavis y
un cóndor. (Discovery Channel). |
Argentavis magnificens
atacando a un Thylacosmilus atrox. (*)
|
|
 |
|
Mariano Magnussen Saffer
del Museo Municipal Punta Hermengo de Miramar, junto al ala
de Argentavis magnificens, recreada para esta
institución a partir de los pocos restos conocidos.
|
Ave.
Un esqueleto parcial de este enorme
teratornítido, fue encontró de la provincia de La Pampa, Argentina.
Es la mayor ave voladora que haya existido. Es también el
teratornítido más antiguo encontrado, ya que data de fines del
Mioceno, entre 6 y 8 millones de ańos, y uno de los escasos
teratornítidos encontrados en América del Sur. El primer
descubrimiento incluyó porciones del cráneo, un húmero incompleto y
varios otros huesos del ala. Incluso los estudios realizados,
estiman su envergadura en por lo menos 6 metros, y podría haber
llegado hasta los 8 metros. Su peso se estima en alrededor de 80
kilogramos. Su peso y su envergadura rivalizan entonces con los de
los más grandes pterosaurios. Por su tamańo, seguro que fue un ave
planeadora, limitando el aleteo al carreteo y posterior despegue,
aprovechando las fuertes corrientes térmicas para su majestuoso
vuelo que provenían del océano Pacifico, antes de que las
cordilleras llegaran a las alturas que presentan en la
actualidad. Por lo tanto, Argentavis magnificens resulto
ser hasta el momento el ave voladora de mayor tamańo del mundo.
Onactornis depressus.
Cabrera, 1936.
|
Onactornis
depressus Onactornis depressus Onactorn |
Onactornis
depressus Onactornis depressus |
|
 |

|
|
Cráneo de Onactornis depressus
del Mioceno tardío de Carohue, Provincia de Buenos
Aires, Argentina. Museo de La Plata. |
Aspecto en vida de Onactornis
depressus. (*) |
Ave. Todos los Paleontólogos se
ponen de acuerdo de que Onactornis depressus fue el
ave corredora más grande de todas las épocas, hasta el
descubrimiento de Kelenken. Su nombre significa "Jefes
de las aves" y no es por nada. Su masa corporal está estimada en
unos 800 kilogramos aproximadamente. Todas las aves corredoras que
se diversificaron en Sudamérica por lo general son carnívoras, por
acepción de este ejemplar, el cual era carrońero, o por lo menos los
investigadores creen esto. Su enorme tamańo y un cuerpo robusto y
grande no reúne las características necesarias para ser un
depredador. Pero seguro que su gigantesco cuerpo asustaba a otros
depredadores como Thylacosmilus, apropiándose
rápidamente de su presa mal herida o muerta. Tenía un pico muy
desarrollado y duro, especializado en romper huesos. Su tamańo era
de aproximadamente de 2,5 metros de altura, y sus alas eran tan
reducidas que prácticamente pasaban de ser percibidas. El desarrollo
de grandes zonas de praderas y altos pastizales, sumados a la falta
de depredadores primarios en el continente, es la explicación que
encuentran los científicos para explicar la magnitud de estos
fabulosos animales desaparecidos, los cuales, ecológicamente
ocuparon los nichos ecológicos vacantes que dejaron los dinosaurios
predadores luego de su extinción. Sus restos provienen
principalmente de los depósitos sedimentarios de la Provincia de
Buenos Aires, Argentina, cuya antigüedad seria de 6 millones de ańos
antes del presente. Otra especie conocida es Onactornis
mendocinus.
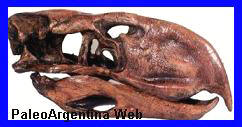
Brontornis
burmeisteri. Moreno & Mercerat, 1891.
|
 |
 |
 |
|
Cráneo y pico. (*) |
Falanges. (*) |
Recontracción. (*) |
Ave. Es
un género extinto de aves no voladoras gigantes y
depredadoras que vivían en la Patagonia. La
única especie actualmente aceptada como válida es B.
burmeisteri, ha sido tradicionalmente incluida
en la familia Phorusrhacidae ("aves del terror") por su
gran tamańo y estilo de vida depredadora, y más
precisamente en la subfamilia Brontornithinae, que
contenía formas muy grandes y corpulentas. Se conoce a
partir de huesos, principalmente de las patas y los
pies, pero también por algunas piezas de cráneo y
columna vertebral, que se encuentra en varias
localidades de la provincia de Santa Cruz. B.
burmeisteri fue la especie más grande de su
grupo, con una altura de alrededor de 2,80 metros y un
peso estimado de 350-400 kilogrmos, por
lo que es el tercera ave más pesada según los
conocimientos actuales (después de Aepyornis
maximus y Dromornis stirtoni), y
los depredadores terrestres más grandes de su tiempo y
lugar. Debido a su volumen, es probable que tuviera un
estilo de vida entre un depredador de emboscada y uno
que perseguía activamente la presa, saltando sobre ella
y derribándola por la fuerza de ataque después de una
corta persecución. Era el carnívoro dominante de la
Patagonia durante el Mioceno, siendo capaz de matar
incluso a grandes animales como ungulado sudamericano
Astrapotherium y el depredador Thylacosmilus (un
marsupial con dientes de sable). Convivió con algunos
forusrácidos más pequeńos y activos como Phorusrhacos;
pero aparentemente se extinguió antes de la aparición
del inmenso Argentavis, la mayor ave
voladora de la que se tiene constancia.
Psilopterus bachmann.
Moreno & Mercerat, 1891.
|
 |
|
 |
|
Cráneo. (*) |
|
Reconstrucción en vivo.
(*) |
Ave. Es un género
extinto de forusrácido ("ave del terror") que vivió a
mediados del Oligoceno hasta finales del Mioceno en
Argentina. Comparado con otros forusrácidos, los
miembros de este género eran relativamente gráciles y
pequeńos, e incluyen las menores especies de aves del
terror: con la cabeza en alto. P. bachmanni alcanzaba
70 - 80 centímetros de altura y pesaba cerca de 5
kilogramos, mientras que los mayores miembros del género
solo alcanzaban 7 kilogramos. Estas aves se parecen a la
actual Cariama o Chuńa (Cariama cristata),
excepto por su constitución más robusta y alas
considerablemente menores. La gran similitud morfológica
entre las garras de las Cariamas y Psilopterus, siendo
ambas afiladas, curvas y comprimidas lateralmente,
pueden indicar que eran usadas para golpear a las
presas. En contraste con otras aves del terror de mayor
tamańo, los investigadores Tonni y Tambussi también han
sugerido que Psilopterus podría usar sus garras para
trepar a los árboles, y podrían incluso volar, cuyas
ideas han sido rechazada en la literatura científica más
reciente. Otras especies conocidas; P. lemoinei (Moreno
& Mercerat, 1891), P. affinis (Ameghino,
1899) y P. colzecus (Tonni & Tambussi,
1988). También se las puede encontrar con la sinonimia
de Pelecyornis (Ameghino, 1891) y Staphylornis (Mercerat,
1897). La más reciente revisión sistemática de
Phorusrhacida,e sitúa a Psilopterus dentro de la
subfamilia Psilopterinae, junto con los géneros Procariama y Paleopsilopterus,
y divide a Psilopterus en cuatro
especies. Psilopterus lemoinei (Moreno &
Mercerat, 1891) es contemporáneo de P. bachmanni y
probablemente llenaba un nicho ecológico muy similar,
aunque P. lemoinei es levemente mayor, con
un peso promedio estimado en 7 kilogramos. La especie
fue definida por una parte de la parte inferior de la
pata (el lectotipo, MLP-162, es el extremo distal de un
tibiotarso), pero unas amplias variedades de materiales
se han referido a este taxón. Dicho material se ha
hallado en algunos sitios en la provincia de Santa Cruz
de Argentina que datan de mediados del Mioceno (Santacruciense).
Las características de diagnóstico incluyen un cráneo
alto y una mandíbula más alta (maxilar), y que la parte
frontal del agujero en frente de los ojos (borde rostral
de la fenestra anteorbital) está menos inclinado.
Algunas diferencias adicionales en el esqueleto fueron
notadas por Sinclair y Farr (1932). Algunas
discrepancias entre varios especímenes han sido
atribuidas a diferencias de edad o sexo, pero el
material actualmente asignado a P. lemonei y P.
bachmanni puede ser reclasificado a nivel de
especie si son reexaminados en profundidad.
Opisthodactylus patagonicus.
Ameghino 1891.
|
 |
|
 |
|
Restos de
Opisthodactylus sp.
Imagen ilustrativa. |
|
Aspecto de
Opisthodactylus
.Reconstrucción en vivo.
Por MarMag.2026 |
Ave.
Rheiformes.
Se trata de un ave del orden
de los Reiformes, nombre común que reciben dos aves sudamericanas
similares al avestruz, emparentada con el género Rhea del
Pleistoceno y Holoceno, es decir, el ńandú actual. Es una especie
extinta de ńandú del Mioceno de Argentina. Se conocen las especies Opisthodactylus
patagonicus (tipo), Opisthodactylus horacioperezi y Opisthodactylus
kirchneri, en homenaje al expresidente Néstor Kirchner, cuyo
material fósil había sido encontrado en la década del 40 por un
naturalista y permaneció en el Museo Florentino y Carlos Ameghino de
Rosario. El género Opisthodactylus es arduamente
conocido en la Formación Santa Cruz, en la Patagonia Argentina. Una
nueva especie de Rhea, Pterocnemia
mesopotámica, del Mioceno tardío de Mesopotamia de Argentina. Los
especímenes provienen de la base de la Formación Ituzaingó, que
brota a lo largo de los acantilados del río Paraná cerca de la
ciudad homónima de la provincia de Entre Ríos. Era un ave esbelta y
de cuerpo pequeńo, de tamańo similar a Rhea o choique
(Pterocnemia pennata) y dieta omnívora.
También se conoce un tarsometástaro aislado, procedente de la
Formación Aisol en la Provincia de Mendoza.
Macranhinga paranensis.
Noriega, 1992.
|
 |
 |
 |
|
Tarsometatarso
de la especie gigante extinguida Macranhinga
(*) |
Barrancos del
Rió Paraná. Portador de fósiles. (*) |
Aspecto de
Biguá-víbora fósil Macranhinga paranensis de mayor tamańo
que su representante viviente. (*) |
Ave. Las anhingas son aves integrantes
del orden de los Pelecaniformes, que habitan ambientes de agua dulce
y tienen estrechos vínculos de parentesco con los cormoranes y los
biguás. Presentan un largo y delgado cuello y un pico recto y muy
puntiagudo. Son buenas voladoras, alternando el vuelo batido de sus
alas con el planeo en las corrientes ascendentes de aire caliente.
La propulsión en el agua es imprescindible para ellas, ya que su
dieta se compone casi exclusivamente de peces. Nadan lentamente
sobre la superficie, con la mayor parte del cuerpo sumergido y
asomando el cuello y la cabeza, lo que les da el aspecto de una
víbora. Su nombre vulgar deriva precisamente de esta apariencia.
Este tipo de natación es alternada con cortos períodos de buceo, en
los cuales "arponean" a los peces con sus afilados picos. A manera
de síntesis simplificada de la historia geológica de la región,
podemos decir que los sedimentos más antiguos de la base de las
barrancas del Paraná, integrantes de la Formación Paraná, fueron
depositados por una extensa ingresión marina que a fines del Mioceno
medio (hace aproximadamente 15 millones de ańos), el cual, formó el
denominado "Mar Paranense". Dicho mar penetró desde el Océano
Atlántico, cubriendo el noreste de la Argentina, oeste del Uruguay y
hasta el sur del Paraguay. Sus aguas templado-cálidas y poco
profundas albergaron una rica fauna de peces óseos, tiburones y
rayas; entre los mamíferos fueron abundantes los cetáceos, como así
también se ha reportado la presencia de restos fósiles de focas y
dugongos o vacas marinas. No obstante, constituyen las evidencias
fósiles más comunes y fácilmente reconocibles, las variadas formas
de invertebrados que habitaban el litoral de aquel antiguo mar,
destacándose entre los moluscos las enormes ostras. Con
posterioridad al retiro del "Mar Paranense" y a lo largo de la
amplia cuenca del incipiente río Paraná, comenzaron a depositarse
desde el Mioceno tardío y también durante el Plioceno
(aproximadamente entre los 10 y los 5 millones de ańos antes del
presente) sedimentos fluviales compuestos principalmente por arenas
blancas, amarillas y ocráceas. Estos sedimentos conforman la porción
mayoritaria del paquete de estratos típicos observables hoy en día
en las barrancas de los alrededores de la Toma Vieja en la ciudad de
Paraná. La base de esta última formación geológica, denominada
Formación Ituzaingó, es portadora de una rica y variada fauna
extinta de vertebrados que se conoce como "Mesopotamiense". Entre
las aves exhumadas de aquella fauna "Mesopotamiense" es
particularmente interesante el hallazgo de restos fósiles
pertenecientes a una especie de biguá-víbora o anhinga gigante, cuyo
nombre científico es Macranhinga paranensis, que
habitó las márgenes forestadas de aquel primitivo río Paraná, y
seguramente tuvo comportamientos muy similares a sus representantes
vivientes.
Reyesichnus punensis.
Alonso et al. 1980.
|
 |
|
 |
|
Imagen
ilustrativa. |
|
Aspecto de
Reyesichnus punensis.
Por MarMag.2026 |
Ave. Los animales que vivieron en el
pasado geológico dejaron pruebas irrefutables de su existencia en
forma de huesos, dientes, conchillas, caparazones o bien por las
impresiones de sus pisadas conservadas como huellas. Dichas huellas
se conocen como icnitas y su campo de estudio es la icnología.
Cuando se trata de huellas fósiles entonces es la paleoicnología. Se
descubrieron abundantes huellas de aves fósiles en la Puna. Las más
antiguas se encontraron en unas lajas rojas de Farallón Catal en el
salar del Hombre Muerto y tenían una edad de 15 millones de ańos.
Corresponden a un representante antiguo de los teros, bautizado
como Reyesichnus punensis, en homenaje al Dr. Celso
Reyes, que fuera profesor de la Universidad Nacional de Salta. En
aquel tiempo la Puna, al igual que ahora, estaba salpicada de lagos
salinos de distintos tamańos. En esos lagos vivía una abundante
avifauna que medraba en sus playas y dejaba impresas sus huellas en
el barro fresco. Hay huellas pequeńas, medianas y grandes;
tridáctilas y tetradáctilas; con o sin membranas interdigitales,
entre otros detalles. Se han identificado representantes
morfológicos de los actuales flamencos, patos, guayatas, teros,
teritos y otros pájaros y zancudas.
Palaeospheniscus bergi.
Moreno y Mercerat, 1891
Ave.
El género Palaeospheniscus fue
nombrado por François Moreno y Alcide
Mercerat en 1891 para designar una serie de
especímenes fósiles procedentes de la
Patagonia argentina. La especie tipo del
género corresponde a Palaeospheniscus
patagonicus, cuyo holotipo consiste
en un tarsometatarso aislado que se
encuentra depositado en el Museo de La
Plata, que proceden de la Formación Gaiman
(Mioceno temprano). Capas portadoras de
fósiles marinos del Cenozoico medio afloran
en cerro Plataforma en la Provincia del
Chubut. Aquí fueron hallados restos de
pingüinos y un diente de tiburón bien
preservados, además se encontraron fósiles
de invertebrados se incluyen corales,
briozoos, abundantes moluscos, equinoideos y
cangrejos. Muchos de los fósiles de
pingüinos hallados en Patagonia han sido
asociados a este género, al punto que en
1946 George Gaylord Simpson mencionaba nueve
especies (la mayoría de las cuales Simpson
consideraba sinónimos de la especie tipo).
El mismo autor, en una revisión posterior
realizada en 1975, solo consideraba validas
cuatro especies. En la actualidad solo se
incluyen tres especies en el género: Palaeospheniscus
patagonicus, Palaeospheniscus
bergi y Palaeospheniscus
biloculata. Igualmente, los géneros
patagónicos Paraspheniscus, Trelewdyptes, Chubutodyptes y Perispheniscus han
sido actualmente incluidos en alguna de
estas tres especies. Los análisis recientes
sugieren que Palaeospheniscus se
encuentra más próximo a los pingüinos
vivientes que a los pingüinos gigantes del
Paleógeno o al género Paraptenodytes.
Seguramente fue un gran nadador y se
alimentaba principalmente de peces. Llego a
medir unos 80 centímetros de altura.
Gryposuchus
neogaeus. Burmeister, 1885.
|
 |
|
 |
|
Cráneo exhibido en el Museo
de La Plata. |
|
Paleoreconstruccion. (*) |
Cocodrilo. Es un
género extinto de crocodiliano gavialoide. Además, es el género tipo
de la subfamilia Gryposuchinae. Sus fósiles han sido hallados en
varios países de Sudamérica, como Argentina, Colombia, Venezuela,
Brasil y Perú. Existió durante principios y mediados del período
Mioceno. Una especie recientemente descrita, Gryposuchus croizati, tiene
una longitud estimada de 10 metros. Especímenes de esta especie
fueron descritos inicialmente de Argentina en 1885, aunque fueron
referidos al género Ramphostoma. No fue sino hasta
1912 que se le asignó su propio género con la descripción de una
nueva especie de Brasil, Gryposuchus jessei. Algunos
gaviales griposuquinos como Siquisiquesuchus y Piscogavialis han
sido hallados en localidades que se cree eran depósitos de ambientes
costeros. La presencia de Gryposuchus en la formación
Urumaco de Venezuela, que incluye estratos marinos, le da crédito a
la idea de que los griposuquinos pueden haber vivido en ambientes
costeros. Sin embargo, ciertas localidades donde se recuperó
material perteneciente a la especie Gryposuchus colombianus,
como la zona de La Venta, Colombia , claramente eran depósitos de
ambientes de agua dulce, lo que indica que la hipótesis de u estilo
de vida costero para los griposuquinos no puede generalizarse. Fue
un activo czador de peces, pequeńas tortugas, anfibios entre otros
animales.
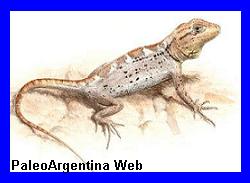
Pristidactylus. Fitzinger,
1843.
 |
|
 |
|
Cráneo y mandíbula de
Pristidactylus sp. |
|
Aspecto de Pristidactylus. Reconstrucción en vivo.
Por MarMag.2026 |
Reptil.
Los iguania es el clado de lagartos más diverso en
número de especies y más ampliamente distribuido en América del Sur,
aunque su registro paleontológico en este territorio es aún muy
escaso en comparación con el de América del Norte y Central. Los
iguanios del Cenozoico sudamericano se registran en el Paleoceno de
Bolivia y Brasil (Estes, 1983; Rage, 1992), Mioceno de Argentina y
Colombia. Conocida como especie tipo Leiosaurus fasciatus (Duméril
y Bibron 1837), se ha encontado su presencia como iguanio en el
Mioceno Temprano de la Patagonia Argentina. Justamente poseen una
antigüedad Terciaria o aún Cretácica para el género. La presencia de
restos de Pristidactylus en el Mioceno Temprano de
Gaiman apoya la hipótesis de un origen al menos Terciario y
fenómenos subsecuentes de vicariancia como respuesta al elevamiento
de la cordillera andina. El paleoambiente propuesto para el
Colhuehuapense de Gaiman, caracterizado por áreas boscosas y el
desarrollo de dunas de arena, proponen un modelo de especiación
eco-geográfica para el género Pristidactylus, se
considera que desde un primitivo ancestro común que habitaba
extensos bosques, en deterioro a través del tiempo, algunas
poblaciones disyuntas quedaron en refugios aislados y adquirieron
pre-adaptaciones a ambientes más abiertos, rocosos, secos y con
formaciones arbustivas, evolucionando hacia los presentes taxones
conocidos en Sudamérica, que se diferencian para distintos hábitats.
Esta hipótesis requiere ser contrastada con un reanálisis de la
filogenia del género basada en caracteres de diversas fuentes e
incluyendo a Pristidactylus nigroiugulus, que no fue
particularmente considerada en la filogenia. Llego a medir unos 12
centimetros de largo. Tenía cabeza alargada, triangular, y su cola
es más larga que el cuerpo. Era principalmente insectívoro, y
completo su dieta mínimamente con vegetales. Pudo ser capturado por
mamíferos carnívoros y aves rapaces.
Callopistes
rionegrensis. Quadros. et
al. 2018.
|
 |
|
 |
| |
|
|
Reptil.
El resto fósil hallado corresponde a un cráneo de 5
centímetros de largo proveniente de los sedimentos de la Formación
Chichinales en los afloramientos del Área Natural Protegida Paso
Córdoba, en las afueras de la ciudad de General Roca, Provincia de
Rio Negro. Constituye uno de los cráneos más completos de un lagarto
fósil de Argentina. Callopistes es un género de
lagartos que actualmente cuenta con dos especies vivientes, Callopistes
maculatus y Callopistes flavipunctatus, que
habitan desde el norte de Chile hasta el sur de Ecuador. Además,
existe un representante fósil de la localidad de Monte Hermoso,
provincia de Buenos Aires, Callopistes bicuspidatus,
que es más reciente que la nueva especie rionegrina ya que la
formación geológica de la localidad bonaerense tiene unos 7 millones
de ańos. Este nuevo hallazgo ubica su aparición hace unos 20
millones de ańos, reforzando hipótesis sobre el origen y la
distribución de estas especies en el sur de Sudamérica y aportando
novedosa información sobre la evolución de todo este grupo de
reptiles. Forman parte de la familia Teiidae, encontrando entre sus
representantes sureńos vivientes al Lagarto Colorado (Salvator
rufescens), el Ututu coludo Rojo (Aurivela longicauda)
y el Teiu (Teius ocelatus). El lagarto Callopistes
rionegrensis tenía unos 50 o 60 centimetros de longitud, que
al igual que sus parientes actuales sería un importante predador de
pequeńos animales.
Boidae.
Gray, 1825.
|
 |
|
 |
|
Vértebra de un gran Boidae. al
lado, vértebras de otra boa gigante del Eoceno de patagonia,
Chubutophis grandis. |
|
Aspecto de la boa gigante del
Mioceno de Patagonia. Reconstrucción en vivo. Por MarMag.2026 |
Ofideo. Los boidos o boas son una
familia de serpientes constrictoras, es decir que matan a sus presas
por constricción, encerrándolas en sus anillos. A diferencia de sus
cercanas parientes, las pitones (Pythonidae), son ovovivíparas. A
ambos lados de la cloaca presentan espolones, que son vestigios de
las patas posteriores locomotoras. Entre las boas se encuentran
algunas de las especies de serpientes más grandes. Pero en el actual
desierto patagónico, hace unos 15 millones de ańos existía una
exuberante selva, semejante a la amazónica, con grandes lagartos,
perezosos, monos y mucho más. Hace tiempo, un Paleontólogo del Museo
Argentino de Ciencias Naturales de Buenos Aires se encontraba
revisando las colecciones de dicha institución, hallo el fragmento
de una enorme vértebra que, en un principio, fue atribuida a restos
de un Dinosaurio. Al consultar los registros del inventario del
Departamento de Paleontología de Vertebrados "Florentino Ameghino"
del Museo porteńo, se dieron cuenta los investigadores de que se
trataba de un enorme vertebrado que vivió durante el Terciario,
mucho después de la desaparición de los grandes reptiles de la Era
Mesozoica. La pieza en cuestión fue hallada por el Geólogo Roberto
Ferello en el ańo 1953, en sedimentos de origen continental de la
zona comprendida entre los lagos Musters y Colhue Huapi, al sur de
la Provincia de Chubut. Hoy sabemos que esta vértebra pertenece a un
Ofidio (Serpientes y víboras) más grande de todos los tiempos.
Calculamos después de hacer varias comparaciones con especies
vivientes y extinguidas, este enorme animal midió unos 20 metros de
largo. Su cráneo tuve que medir unos 70 centímetros, con lo que la
abertura bucal le permitía engullir presas de más de un metro de
circunferencia, como los astrapotherios y otros notoungulados de la
época.
Gaimanophis tenuis. Albino,
1996.
|
 |
|
 |
| Vértebra de
Gaimanophis powelli,
del Mioceno de Tucumán. |
|
Aspecto de la boa de Patagonia y NOA,
Gaimanophis powelli. Reconstrucción en vivo.
Por MarMag.2026 |
Ofideo. Es un género boido extinto,
hallado en el acantilado sur del valle inferior del río Chubut,
cerca de la localidad de Gaiman. Los climas del Paleoceno y Eoceno
se caracterizaron por condiciones tropicales bastantes cálidas y se
extendieron de norte a sur hacia los polos. En este escenario, las
grandes serpientes eran relativamente diversas en Sudamérica. En
Patagonia se hallaron formas pequeńas, como Waincophis,
medianas (Boa) y grandes como Chubutophis, pero solo
los géneros pequeńos Waincophis y Gaimanophis,
persistieron en el Mioceno. La ausencia de serpientes grandes en el
Mioceno patagónico con respecto a las de períodos anteriores se ha
relacionado con la disminución de las temperaturas, lo que es
consistente con los cambios conocidos en las temperaturas medias
globales. Gaimanophis tenuis, es conocida por
vértebras aisladas del Mioceno temprano de la Patagonia.
Recientemente se dio a conocer Gaimanophis powelli, de
mayor tamańo, procedente de la Formación India Muerta (Mioceno
tardío) en el Dique El Cadillal de la provincia de Tucumán, en el
del noroeste de Argentina. Chubutophis, fue
una serpiente constrictora, es decir que matan a sus presas por
constricción, encerrándolas entre sus anillos. Posiblemente vivía en
madrigueras abandonadas por otros animales o en huecos de árboles.
|
Caiman lutescens. Rovereto
1912.
|
|
Caimán latescens
Caimán latescens Caimán latescens Caimán latescens Caimán
latescens Caimán latescens Caimán latescens |
|

|

|
|
|
Placas dermicas óseas de
Caimán latescens y Caimán yacare, recuperados del
Plioceno de Entre Ríos. Museo de La Plata. |
Vista superior de cráneo de
Caimán sp. |
Aspecto del genero de cocodrilo
prehistórico Caimán latescens.
Reconstrucción en vivo. Por MarMag.2026 |
Reptil.
Los cocodrilos constituyen un grupo originado en el
Triásico. Su posterior desarrollo y expansión ocurre desde el
Cretácico y Terciario, especialmente asociado con regiones de
latitudes medias-tropicales a ecuatoriales. Han ocupado un nicho
ecológico amplio, tanto marino como continental, siempre vinculado
con áreas acuáticas (costaneras, humedales, etc.). Los cocodrilos no
son buenos indicadores paleoecológicos, aunque su presencia está
relacionada a climas cálidos en lugares donde el agua no escasee.
Las dos especies actuales de caimanes en el país viven asociadas a
cuerpos permanentes de agua. La mayor parte de los materiales
recolectados proceden de los afloramientos de la Formación Ituzaingó,
a unos 800 metros al sur del puerto de Villa Urquiza. En la zona del
puerto y hasta unos 400 metros al norte y sur del mismo, aflora un
paquete de unos 6 metros de arcillas verdosas y arenas blanquecinas
o gris verdosas que contienen niveles con Ostrea patagonica y Pecten
paranensis (invertebrados). Este nivel pertenece a la
Formación Paraná (Mioceno medio) y es recubierto en discordancia por
las arenas blanquecinas y amarillentas de la Formación Ituzaingó
(Mioceno superior - Plioceno). Esta última unidad estratigráfica, de
origen fluvial, labró paleocauces sobre la marina precedente
produciendo en su contacto acumulación de material clástico más
grueso y, particularmente, una concentración de material óseo de
diverso origen. Estos restos, al igual de los que fueron
anteriormente descriptos provienen de un nivel estratigráfico
inferior de la Formación Ituzaingó, al cual, informalmente, se lo
conoce con el nombre de «mesopotamiense» y que representa niveles de
mayor energía en un ámbito fluvial desarrollado sobre la secuencia
marina de la Formación Paraná. Esto plantea la posibilidad que
algunos de estos cocodrilos hayan vivido en el ámbito marino, otros
en los costaneros y en humedales. Anteriormente se han
colectado huesos fósiles de cocodrilos del ‘Mesopotamiense’, entre
ellos el dentario derecho de un alligatórido gigante, al cual
denomina Alligator lutescens, sobre el cual se discute
su sinonimia con Caiman tutescens.
Chelonoidis
gringorum.
Simpson, 1942.
|
 |
|
 |
| Fósil del caparazón de
Chelonoidis gringorum en el MEF.
|
|
Posible aspecto de
Chelonoidis gringorum.
Reconstrucción en vivo. Por MarMag.2026 |
Reptil. En el Mioceno Inferior
de Patagonia fue hallada una tortuga de gran tamańo, llamada Chelonoidis
gringorum probablemente precursora de Chelonoidis
chilensis, también fósil en los depósitos pleistocénicos de
Córdoba. Otros dos taxa, muy cercanos a Chelonoidis chilensis, Chelonoidis
donosobarrosi y Chelonoidis petersi (vivientes).
Rasgo sobresaliente del orden Testudines, es la presencia de una
envoltura externa de consistencia variable, vulgarmente conocida
como caparazón o concha, en íntimo contacto con el eje vertebral del
esqueleto y constituida por una porción ósea profunda recubierta por
láminas o placas córneas dérmicas. Integran el caparazón,
dorsalmente el denominado carapax, ventralmente el plastrón, unidos
lateralmente por una conexión, o puente, y ambos formados por huesos
aplanados, articulados entre ellos mediante suturas apareciendo
proyectados hacia el exterior los marginales, como aleros de un
tejado. Resulta así asegurada la notoria solidez estructural del
caparazón. Estas tortugas marcan su permanente presencia en áreas
relativamente boscosas, cerca de algarrobos, grandes cactus, etc,
determinando restringidos espacios de abrigo casi libre de
vegetación. La alimentación estária constituida por frutos y hojas
de cactáceas, cucurbitáceas, leguminosas, gramineas, etc. Fósiles
de Chelonoidis gringorum provienen de afloramientos
Terciarios de la Formación Sarmiento expuestos en la ribera sur del
valle del río Chubut, entre Gaiman y Dolavon (Provincia de Chubut,
Argentina). Otras tortugas del Mioceno tuvieron amplia dispersión,
con formas de robustas dimensiones como Geochelone (Chelonoidis)
gallardoi en Catamarca.
|
Phrynops geoffroanus.
Schweigger, 1812. |
|
Phrynops
geoffroanus Phrynops geoffroanus Phrynops geoffroanus
Phrynops geoffroanus Phrynops geoffroanus Phrynops |
|
 |

|

|
|
Ejemplar de fósil de
tortuga. Ilustrativo. |
Placas del caparazón de
Phrynops,
de la prov. de Entre Ríos. Museo de La Plata. |
Aspecto del Quelonio
Mioceno Phrynops geoffroanus. Reconstrucción en vivo.
Por MarMag.2026 |
Reptil. El registro fósil de
tortugas en América austral es muy rico e incluye localidades en el
sur de la Patagonia donde las condiciones climáticas actuales
impiden el establecimiento de quelonofauna. Los fósiles más
antiguos que se conocen de la familia Chelidae datan del Cretácico
inferior en Patagonia. Estos quélidos dulceacuícolas actualmente
están representados por algunas especies vivientes con una
distribución disyunta en América del Sur, desde el norte de
Venezuela hasta la Argentina central templada y Australasia, como
resultado de un pasado biogeográfico gondwánico. El género Phrynops está
registrado desde el Mio-Plioceno de la Argentina y el Mioceno
superior de Uruguay. Los registros fósiles atribuidos al género Phrynops,
por el momento están restringidos a sedimentos del noroeste
argentino (Catamarca y Tucumán) y al área de la Mesopotamia, en
Argentina; y al Departamento de San José en Uruguay. Restos fósiles
han sido asignados a Phrynops también se mencionan para
sedimentos pleistocenos del Carcarańá, provincia de Santa Fe. Desde
el siglo pasado, la existencia de placas fósiles y material
fragmentado ya ha sido mencionada por Bravard (1858); Burmeister
(1885); Ambrosetti (1893) quienes determinaron a nivel específico
placas aisladas, pese a la naturaleza fragmentaria del material. Wieland
(1923); cita para la región del Paraná a Parahydraspis
paranaensis como un nuevo taxón dentro de pleurodira.
Mlynarski al referirse a las tortugas de Paraná, consideró a Parahydraspis
paranaensis como un sinónimo de Phrynops geoffroanus. En
el "Mesopotamiense" se han registrado abundantes y numerosas
tortugas, principalmente perteneciente a la familia Chelidae,
género Phrynops. Los quélidos, considerados buenos indicadores
paleoambientales, seńalarían condiciones subtropicales y húmedas, en
áreas bajas, inundables y con abundante vegetación. Por lo cual no
resulta raro que haya existido una mayor variedad de tortugas
acuáticas, entre otros reptiles.
Necrolestes patagonensis.
Ameghino,
1891.
|
 |
 |
 |
|
Rama
mandibular sobre un dedo humano. Fuente Ciencia Hoy. |
Cráneo y mandíbula de
Necrolestes
patagonensis
(*): |
Posible aspecto de
Necrolestes. (*) |
Mamífero. Es un enigmático género de
mamífero extinto, de difícil clasificación taxonómica, que vivió a
principios del Mioceno de Sudamérica. Cerca de un tercio del
esqueleto del animal, incluyendo la mayor parte del cráneo, ha sido
hallado con huesos desarticulados de varios individuos. La mandíbula
se curva hacia la punta, posiblemente para sostener un apéndice
carnoso similar a los tentáculos sensitivos del topo nariz de
estrella. Necrolestes es asimismo reconstruido como
una criatura parecida a un topo. Probablemente se alimentaba de
insectos o lombrices. La especie tipo, Necrolestes
patagonensis, fue nombrada y descrita por el conocido
naturalista argentino Florentino Ameghino en 1891, a partir de
restos de edad Santacrucense, que comprendían dientes y un resto
izquierdo de mandíbula inferior, considerándolo como de algún género
de mamifero Insectivoro. Una segunda especie, Necrolestes
mirabilis ha sido descrita del Mioceno argentino, en la zona
de Gran Barranca (Barranca sur del lago Colhue Huapi), Departamento
de Sarmiento, provincia del Chubut, y pese a ser más algo más
antigua que la especie tipo (de edad Colhueapense), es algo mayor y
parece ser tan especializada como Necrolestes
patagonensis, según se desprende del análisis de sus restos
dentales. Su clasificación aún no está bien resuelta, debido a que
era altamente apomórfico y tenía una anatomía distinta a cualquier
otro mamífero conocido, vivo o extinto. Es posible que pertenezca al
mismo linaje que los marsupiales, Metatheria. Por otro lado, la
posibilidad de que fuera un euterio no puede ser excluida basándose
en los datos actualmente disponibles, dado que, cuando era un
continente independiente, Sudamérica tuvo varios linajes endémicos
de mamíferos tanto marsupiales como placentarios. Un análisis
filogenético realizado por Guillermo Rougier et al. (2012) encontró
que Necrolestes tiene una inesperada posición
filogenética como un mamífero no terio que pertenecía al clado
Meridiolestida y estaba particularmente relacionado de cerca con los
géneros Cronopio y Leonardus; si esto se
confirma convertiría a Necrolestes en el miembro más
reciente conocido de este grupo. Los propios meridioléstidos fueron
clasificados en principio como miembros del clado Dryolestida, pero
el análisis de Rougier et al. (2012) los encuentra como algo más
cercanamente relacionados con los mamíferos placentarios,
marsupiales y los anfitéridos que a los miembros de Dryolestida.
Lycopsis torresi.
Cabrera, 1927
|
 |
 |
|
Parte del esqueleto puesto en forma
ilustrativa de ejemplar colombiano. |
Reconstrucción en vivo.
Por MarMag.2026 |
Mamífero Marsupial.
Es un género extinto de mamífero
carnívoro metaterio del orden Sparassodonta, emparentado de cerca
con los actuales marsupiales. Hasta la actualidad se han descrito
tres especies, Lycopsis torresi del Mioceno de Santa
Cruz en Argentina, Lycopsis viverensis que vivió
durante el Chasiquense de la provincia de Buenos Aires, y Lycopsis
longirostrus. Esta última especie es la mejor conocida a
mediados del Mioceno. Un esqueleto casi completo ha permitido
mostrar que era un animal de contextura grácil y talla reducida, de
1,1 metros de longitud, unos 35 centímetros de altura hasta los
hombros y aproximadamente 15 kilos de peso, dotado de un hocico
alargado que le daba un perfil vagamente similar al de los cánidos.
Probablemente se trataba de una especie que cazaba a presas de
tamańo reducido (se encontraron restos del roedor Scleromys
colombianus en la región del vientre del esqueleto de un
ejemplar de Lycopsis longirostrus) a través de la
emboscada en el medio boscoso como lo fue el área de La Venta
(Colombia) durante el Mioceno, lo que bien pudo haberle evitado
competencia de otros carnívoros mayores.
Microtragelus argentinus.
Ameghino, 1904.
|
 |
|
 |
|
Recreación del esqueleto de un Microtragelus a partir
de los fósiles conocidos. Museo de Ciencias Naturales de Miramar. |
|
Posible
aspecto
Microtragelus.
Ambas
recreaciones realizadas por Mariano Magnussen. |
Mamífero Marsupial.
Fue un pequeńo mamífero marsupial de
tamańo pequeńo, como un ratón, de aproximadamente 40 centímetros de
largo, emparentado con los actuales canguros de Australia y las
zarigüeyas de América del sur. Su apariencia es muy semejante a las
actuales ratas canguros de Egipto. Tenía largas patas traseras y
brazos cortos. Sus mandíbulas eran con apariencia rodariforme. Tal
vez, tuvo orejas largas, pero estas no se han preservado como
fósiles. Se alimentaba de vegetales e insectos, y probablemente tuvo
hábitos crepusculares o nocturnos. Sin embargo, Microtragelus fue
parte de una evolución notable e independiente que tuvo lugar en
Sudamérica durante los últimos 70 millones de ańos, cuando esta
quedara aislada del resto de las masas continentales y se
desarrollara una fauna única y que no se repitió en otras partes del
mundo. Estos marsupiales, mamíferos con bolsa, habían ocupado nichos
ecológicos que en otras partes del mundo estaban ocupados por
mamíferos placentarios. Otras especies conocidas; Microtragelus
bolivianus, Microtragelus
catamarcensis y Microtragelus
reigi.
Borhyaena macrodonta.
Ameghino, 1897.
Mamífero
Marsupial.
Era parecido a un oso, con las patas
planas. Fue un marsupial carnívoro del orden Sparassodonta, del
tamańo de un lobo, de 1,5 metros de longitud. Eran depredadores muy
importantes durante el Mioceno de la Patagonia Argentina, los cuales
se alimentaban de mamíferos notoungulados y armadillos. Las patas
cortas seńalan que no era un animal corredor, pero seguro que
asechaba a sus víctimas en los antiguos bosques que existían hace 20
millones de ańos en los actuales desiertos de la provincia de Chubut.
Prothylacinus patagonicus.
Ameghino 1891.
|
 |
 |
|
Rama mandibular de
Prothylacinus patagonicus. Tomado de Flickr. (2010) En
el Museo Argentino de Cs Naturales. |
Aspecto de Prothylacinus
patagonicus. Por
MarMag.2026. |
Mamífero Marsupial. Este es un
mamífero marsupial extinto, similar a un perro. Vivió en el Mioceno
inferior (hace unos 18 millones de ańos) de la provincia de Santa
Cruz, en la Patagonia argentina. Este animal tenía un físico
robusto, y su peso fue de cerca de 40 kilogramos. El cuerpo era
largo y flexible, mientras que la musculatura del cuello era muy
poderosa. El cráneo estaba equipado con dientes fuertes, típicos de
un animal carnívoro. Las patas cortas y fuertes estaban equipadas
con potentes músculos, y probablemente con una especie de pulgar
semioponible. Las patas delanteras tenían garras fuertemente
curvadas, mientras que las traseras tenían dedos cortos,
plantígrados, con la planta del pie alargada, probablemente capaz de
captar las superficies curvas. La cola era larga (compuesta por
20-30 vértebras) y muy puntiaguda. Las características del
protilacino sugieren que era un animal arborícola, poderoso y ágil.
Las patas delanteras las utilizaban para sujetarse a las ramas,
mientras que el cráneo y el cuello eran fuertes para poder soportar
el peso de la presa, que capturaba con sus poderosas mandíbulas. Es
posible que el protilacino se alimentase de roedores, marsupiales, y
pequeńos perezosos. El protilacino es el representante más típico de
los Prothylacynidae, del orden Sparassodonta, el cual se desarrolló
en América del Sur ocupando los nichos ecológicos que fueron
ocupados en otros continentes por los mamíferos del orden carnívora.
Arctodictis munizi.
Mercerat, 1891.
|
 |
 |
 |
| Cráneo y
Mandíbula de
Arctodictis munizi. Tomado de Ciencia Hoy.
V22,N130. |
Fósiles de Arctodictis sinclairi en el
Geoparque Bryn Gwyn de
Gaiman, Chubut. |
Aspecto
característico de los
esparasodontos como Arctodictis munizi.
Por MarMag.2026 |
Mamífero Marsupial. Fue un mamífero metaterio
carnívoro originario de América del Sur. Fueron considerados alguna
vez como parte de los marsupiales, pero la evidencia indica que en
realidad son más bien un taxón estrechamente emparentado con estos,
pero independiente. Se parecían mucho a ciertos depredadores
placentarios que evolucionaron separadamente en otros continentes, y
son citados con frecuencia como ejemplos de evolución convergente.
El grupo de los esparasodonte fue descrito originalmente por el
paleontólogo argentino Florentino Ameghino, a partir de fósiles de
los lechos de Santa Cruz en la Patagonia. Los esparasodontos
estuvieron presentes durante buena parte del largo período de
"espléndido aislamiento" de Sudamérica durante el Cenozoico. Durante
este tiempo compartieron los nichos de los grandes depredadores de
sangre caliente, o con las llamadas aves del terror. Junto con los
tilacoleónidos de Australia, Arctodictis munizi fue
uno de los mayores depredadores entre los metaterios. Se caracteriza
por una serie dentales que distinguen al grupo de otros mamíferos
cercanamente relacionados. El inicio de la historia de los
esparasodontos es muy poco conocido, ya que muchos de los miembros
de este grupo del Paleoceno y el Eoceno son conocidos apenas de
dientes aislados y mandíbulas fragmentarias. Otra especie
relacionada; Arctodictis sinclairi.
Patagosmilus
goini.
Forasiepi y Carlini, 2010.
|
 |
|
 |
|
Fragmento de cráneo con
molares y canino de
Patagosmilus goini. Colección Museo de La
Plata. (*) |
|
Aspecto de
Patagosmilus goini,
cazando un
Huayqueriana
(*). |
Mamífero Marsupial. Es un género
extinto de mamífero metaterio de la familia Thylacosmilidae que
vivió a mediados del Mioceno en América del Sur. Al igual que otros
representantes de esta familia, como Thylacosmilus atrox o Anachlysictis
gracilis, se caracterizaba por sus enormes colmillos en la
mandíbula superior, similares a los conocidos felinos "dientes de
sable" (Machairodontinae), de los que eran equivalentes ecológicos
sin parentesco algunos. La morfología general de Patagosmilus sugiere
que era anatómicamente menos especializado que Thylacosmilus del
Plioceno, pero la morfología de los dientes, sin embargo, sugiere
que probablemente estaba más estrechamente relacionado con este
último, que con el más primitivo Anachlysictis de
Colombia. La especie Patagosmilus goini, que debe su
nombre al paleontólogo argentino Francisco Goín, especialista en
marsupiales, fue descrita a partir de los restos de un cráneo
aplastado y parte de una falange ungual que se descubrió en
sedimentos que datan del Mioceno Medio en la orilla oeste del Río
Chico, en la provincia de Río Negro en la Patagonia, Argentina y
depositados en el Museo de La Plata. Este es el primer representante
de Thylacosmilidae del que se han encontrado restos en la Patagonia,
y el primer género reconocido aparte de Thylacosmilus y Anachlysictis que
es un miembro indiscutido de esta familia. Seguramente fue un
importante marsupial depredador.
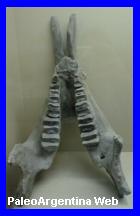
Astrapotherium magnum.
Owen 1853.
|
|
|
|
|

|
|

|
|
Cráneo y
mandíbula de Astrapotherium magnun en el Museo Egidio
Feruglio (MEF). |
Mandíbula
inferior. Museo de La Plata. |
Reconstrucción
en vivo del extrańo Astrapotherium magnun. Imagen
Sergey Krasovskiy. |
Mamífero
Astrapoterio.
Mientras los Marsupiales evolucionaban, los
Notoungulados hacían lo mismo para colonizar todos los ambientes. Astrapotherium es
un género extinto de mamíferos placentarios del orden Astrapotheria,
superorden Meridiungulata propio de Sudamérica. Eran del tamańo de
un rinoceronte y probablemente tenían una trompa corta y flexible.
Su nombre significa "animal estrella" o "bestia del rayo". El
astrapoterio mejor conocido es Astrapotherium magnum del
periodo Mioceno Inferior-Medio, cuyos restos provienen de las rocas
denominadas Formación Santa Cruz en Argentina, que tienen unos 15
millones de ańos. Gracias a que estaban atrapados en cenizas
volcánicas, ha sido posible reconstruir varios esqueletos completos
de astrapotéridos. Un esqueleto casi completo de este animal se
encuentra en el Museo MEF de Trelew. La preservación de la columna
vertebral casi completa y articulada, hizo imposible extraerla con
seguridad. De este modo el ejemplar fue montado con las extremidades
flexionadas, como si estuviera echado sobre su vientre. El esqueleto
permite apreciar que los astrapoterios eran criaturas bastante
distintas como ninguna otra. Astrapotherium era un
animal grande, quizás tanto como un búfalo africano de unos 800
Kilogrmos, o un rinoceronte indio de más de 1000 Kilogramos. Su
cuerpo era largo, de cerca de 2,7 metros de longitud y una altura a
la cruz de 1,40 metros, con patas relativamente cortas, muy similar
en proporciones a un hipopótamo. El cráneo de Astrapotherium era
corto pero la mandíbula inferior era larga, los caninos eran largos
y los molares macizos; los molares de los astrapoterios recuerdan
vagamente a los de los rinocerontes. Otras especies conocidas; A.
angustidens, A. columnatum, A. delimitatum, A. giganteum, A. magnum y A.
nahum.
Homalodotherium cunninghanni.
Lydekker, 1894.
|
 |
 |
 |
|
Maxilar de Homalodotherium
cunninghanni del Mioceno de Santa Cruz, en el Museo de La
Plata. |
Homalodotherium cunninghami en
el Field Museum of Natural History (*). |
Reconstrucción.
Kelly Taylor Illustration. |
Mamífero
Notoungulado.
Es un género extinto de mamíferos placentarios del
orden Notoungulata que vivió en el Mioceno Inferior, hace alrededor
de 20 millones de ańos en Argentina. Los calicoterios
(perisodáctilos de Eurasia), los perezosos terrestres y los osos
panda son un notable ejemplo de convergencia evolutiva con los
homalodoterios. Muchos notungulados no eran mayores que conejos,
pero algunos se parecían más a caballos o a hipopótamos. La mayoría
de los notungulados tenía pezuńas, pero Homalodotherium poseía
garras en los cuatro dedos de cada pata. En América del Sur
evolucionó una amplia gama de animales herbívoros con pezuńas, en
completo aislamiento porque, en aquella época, América del Sur era
una isla-continente como Oceanía. Al contrario que su pariente más
reciente, el Toxodon, los caninos no estaban
desarrollados. Por la forma del cráneo se supone que desarrolló una
pequeńa trompa. Sus extremidades anteriores eran largas, y las
posteriores, más cortas, con lo que abandonaron la posición
cuadrúpeda. En las patas delanteras tenían garras enormes y
curvadas, que probablemente usaban para agarrar las hojas de los
árboles ya que eran ramoneadores en los bosques y es posible que se
pusiesen de pie sobre sus patas traseras para alcanzar las ramas más
altas. Sus miembros eran gruesos, por lo que no parece que pudiera
alcanzar gran velocidad. Medía 2 metros de longitud. La función de
las garras se mantiene en el misterio; quizás usaba las garras para
desenterrar raíces y bulbos jugosos o para agarrarse y trepar por el
tronco de los árboles. Tal vez se incorporaba también sobre las
patas traseras a fin de doblar ramas y comer hojas. Se ha comparado
a los homalodoterios con caballos con garras. La comparación sin
embargo no es demasiado apropiada. Si bien la cabeza y el cuerpo se
parecían un poco a los del caballo, las extremidades eran pesadas y
no servían para correr.
Calchaquitherium mixtum.
Nasif, Musalem y
Cerdeńo, 2010.
|
 |
 |
|
Diente de un Toxodonta.
Ilustrativo. |
Ilustración del
paleoartista. (*): |
Mamífero
Notoungulado.
Los toxodóntidos, fue un grupo de ungulados nativos
que incluye formas de mediano y gran tamańo. Se hallan relativamente
bien representados en sedimentos neógenos de América del Sur. Un
nuevo género y especie de Toxodontidae del Huayqueriense (Mioceno
tardío) de El Cajón Valle en la provincia de Catamarca, fue dado a
conocer. El nombre del grupo deriva del género Toxodon (Pleistoceno),
el primer ejemplar que se descubrió, fue un gran herbívoro del
tamańo del actual rinoceronte negro con tres dedos en cada pata. Los
mamíferos originarios de Sudamérica, evolucionaron
independientemente en dicha isla-continente. Alrededor del ańo 1900,
Florentino Ameghino inició un catálogo que contiene clasificaciones,
estudios, comparaciones y descripciones de más de 9.000 animales
extinguidos, muchos de ellos descubiertos por él. Los notungulados
incluyen más de 100 géneros agrupados en cuatro subórdenes. Los
ungulados sudamericanos proporcionan un caso de aislamiento
geográfico tan notable como el de los marsupiales en Oceanía. Los
meridiungulados se conocen a menudo como los ungulados
sudamericanos. La radiación adaptativa de los ungulados Paleocenos
de América del Sur fue precoz y rápida, lo que dificulta encontrar
un modelo filogenético lo suficientemente explicativo. Procedentes
de pequeńos y primitivos animales herbívoros sin competidores,
evolucionaron independientemente y aislados del resto de los
ungulados durante casi todo el extenso periodo del Cenozoico. Los
toxodontos alcanzan el tamańo de un paquidermo y probablemente
compartían sus hábitos. Particularmente en el Mioceno
tardío-Plioceno de Catamarca se reconocen al menos cuatro especies
agrupadas en tres géneros: Xotodon cristatus Moreno y
Mercerat, 1891, Xotodon cf. X. foricurvatus Ameghino,
1885 y Toxodontherium andinum Kraglievich.
Chasicotherium rothi.
Ameghino, 1887.
|
Chasicotherium
Chasicotherium Chasicotherium Chasicotherium |
Chasicotherium
Chasicotherium Chasicotherium |
|

|

|
|
Cráneo de
Chasicotherium exhibido en el Museo de Ciencias Naturales
de La Plata.
|
Aspecto de
Chasicotherium(*). |
Mamífero
Notoungulado.
Fue un Notoungulado o ungulado sudamericano de gran
tamańo descubierto en la formación Chasico, en la Provincia de
Buenos Aires, cuyos sedimentos poseen una antigüedad entre 10 y 9
millones de ańos. Era un herbívoro que prefería ambientes secos y
abiertos. Guarda al igual que el Toxodon y el Trigodon,
cierta semejanza con los hipopótamos y rinocerontes sin tener
parentesco alguno. Este fenómeno es conocido como "convergencia
adaptativa o evolución paralela", es decir, especies que nunca
tuvieron contacto entre sí, su semejanza es el resultado de
adaptarse a ambientes muy similares y cubrir nichos ecológicos
iguales. Lo más llamativo de Trigodon, es que, en vez
de tener falanges ungeales o pezuńas en sus patas, poseía unas
robustas garras. Su peso fue de una tonelada y media. Era un animal
de gran tamańo, el más grande entre los homalodotéridos, así como su
representante más reciente. Se basa en un cráneo incompleto con la
mandíbula, que se distingue por su reducción en la fórmula dental,
con un canino grande con un cíngulo que corría hacia arriba.
Probablemente estaba más cercanamente emparentado con Homalodotherium que
con otros miembros de la familia, como Asmodeus.
Protypotherium australe.
Ameghino, 1885.
|
 |
 |
 |
|
Restos
esqueletarios de Protypotherium, del Mioceno inferior de
Patagonia, exhibido en el Museo de La Plata.
(*) |
Cráneo y mandíbula
completa de
Protypotherium colectado en Santa Cruz en 1897. |
Reconstrucción.(*) |
Mamífero
Notoungulado.
Los antecesores de Protypotherium son
conocidos desde mediados del Paleoceno en América del Sur, pero el
género propiamente dicho, se lo conoce desde las primeras etapas del
Mioceno. Esta tenía el tamańo algo mayor al de un conejo, de
aproximadamente unos 40 centímetros de largo. Su cola y patas eran
relativamente largas. Su cráneo semejante de un roedor en forma de punta
desde atrás hacia delante. La rareza principal que encuentran los
Paleobiologos en esta especie, se halla en sus mandíbulas, compuestas
por unos 44 dientes sin especialización alguna. Su cuello era corto y su
cuerpo alargado. En sus patas se pueden observar fácilmente las falanges
unguales (base de la garra), que podrían utilizarse para cavar sus
propias madrigueras, o bien apropiarse de las cuevas de otros animales y
modificarlas según sus necesidades. Es probable que Protypotherium comiera vegetales,
brotes, fruto, raíces, flores, etc se alimentasen también de carrońa,
como hacen las liebres actuales y los roedores que viven en las
praderas. Las zarpas muestran claramente robustos dedos con garras, el
origen de las pezuńas. Gracias a los cuales, podrían escavar una
madriguera o ampliar las abandonadas por otros animales. Géneros
relacionados: Protypotherium antiquum, Typotheriopsis chasicoensis,
Pseudotypotherium histatum y Mesotherium cristatum.
Adinotherium ovinum. Owen, 1853.
|
 |
 |
|
Cráneo. (*) |
Reconstrucción. (*) |
Mamífero
Notoungulado.
Es un género extinto de mamíferos notoungulados de
la familia Toxodontidae, que vivió en el Mioceno. Hubo varias
especies similares, con diferencias en el tamańo y otras
características y se asemejaban a una versión más pequeńa de su
pariente Toxodon. Así, tenían una apariencia semejante
a los actuales rinocerontes, aunque de mucha menor envergadura. Sus
patas delanteras eran algo más altas que las de su primo, haciendo
que la altura del hombro y de la cadera fueran iguales y facilitando
la carrera. Se trataría de animales que vivían solitarios, en pareja
o en pequeńos grupos familiares en el caso de las hembras y sus
crías, una por parto. En algunas especies la bóveda craneana está
reforzada, quizá para disminuir los efectos de impactos, durante las
peleas en la época de celo. La alimentación era puramente
vegetariana, y al no ser especies de gran tamańo, podía realizarse
por la noche, para evitar depredadores. Se alimentaban de hojas,
ramas y brotes de plantas. Algunas especies se habrían adaptado a
las llanuras y praderas, siendo más común el abrigo de las selvas y
los bosques de ribera y zonas arbustivas. Se conocen varias especies
de Adinotherium, como Adinotherium ferum,
etc. La especie más común de Adinotherium fue Adinotherium
ovinum. Este animal llegaba a medir 1,50 m de longitud.
Tenía un pequeńo cuerno en lo alto del cráneo que podría jugar su
papel en la temporada de apareamiento. Nesodon es un
animal muy estrechamente emparentado Adinotherium y
además coetáneo suyo, aunque alcanzó mayores dimensiones.
Sinónimos: Phobereotherium Ameghino, 1887 y Noadinotherium Ameghino,
1907.
Typotheriopsis chasicoensis.
Cabrera y
Kraglievich, 1931.
|
 |
|
 |
|
Cráneo de
Typotheriopsis sp. del Mioceno de la
Prov. de Buenos Aires. Museo de La Plata. |
|
Posible aspecto de
Typotheriopsis (*). |
Mamífero
Notoungulado.
Los dientes de Typotheriopsis presentan una
especialización primitiva a comparación de otros notoungulados de la
época. Se asemejaban a los de un roedor, con incisivos promitentes
en la parte anterior, separados por un diastema de los premolares y
los molares situados en la parte posterior. Sin embargo, los
incisivos cincelados, provistos de raíces y que no crecían de forma
permanente como el de los roedores, estabas especializados para
morder y no para roer. Su cráneo era corto y tosco. Su cuerpo era
elastiazado con extremidades relativamente alargadas. Su
alimentación consistía principalmente de los típicos pastizales
pampasicos de la región, con raíces y bulbos que incorporaba a su
dieta.
ichhutherium wayra. Armella, M. A., et
al, 2025.
|
 |
|
 |
| |
|
|
Mamífero
Notoungulado.
Fue un herbívoro de la familia
de los mesoterinos, cuyos restos fósiles fueron hallados a una
altitud de 3900 metros, en la actual puna catamarqueńa.
nombre en quechua significa "viento", es
una prueba de que ciertos mamíferos lograron adaptarse a condiciones
extremas en zonas montańosas. La altitud del descubrimiento sugiere
a los investigadores que estos animales desarrollaron estrategias
para sobrevivir en ambientes fríos y de baja presión de oxígeno.Los
fósiles de mamíferos de esta época son difíciles de encontrar, lo
que hace que este descubrimiento sea aún más relevante para la
reconstrucción de los ecosistemas sudamericanos prehistóricos.
El fósil corresponde al linaje más antiguo de los
mesoterinos, una subfamilia entre los mesotéridos, un grupo de
ungulados nativos sudamericanos que se caracterizan por sus
adaptaciones morfológicas.
El ejemplar, denominado
ichhutherium wayra, presenta rasgos morfológicos con un
carpincho y un wombat (marsupiales australianos). Corresponde a una forma parte de esa fauna única de
Sudamérica que se desarrolló durante 30 o 40 millones de ańos de
forma aislada en Sudamérica. Era un animal herbívoro que pesaba
entre 12 y 15 kilogramos y se alimentaba de pasturas. Su estudio
permitirá a los científicos reconstruir el paisaje y la comunidad
biológica de la Puna durante el mioceno temprano. A través de esta
investigación, los paleontólogos esperan obtener una imagen más
clara de cómo era el ecosistema en aquel entonces.
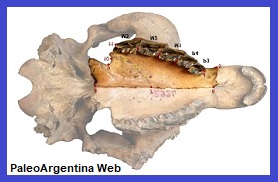
Hegetotherium mirabile.
Ameghino, 1887.
|
 |
|
 |
|
Hegetotherium sp.
Cráneo y mandíbula del Mioceno de
Santa Cruz. Patagonia Argentina.
(*) |
|
Aspecto de
Hegetotherium mirabile.
Por MarMag.2026 |
Mamífero
Notoungulado.
Es un mamífero de la Formación Mioceno Inferior
Santa Cruz de la Patagonia, Argentina. Este mamífero extinto era un
animal semejante a los conejos por su forma, sistema de locomoción y
modo de vida. Poseían patas traseras largas, que les habrán
permitido correr dando grandes pasos, como es característico también
en los lagomorfos y animales similares, como las vizcachas y maras.
Eran importantes herbívoros. Los hegetotéridos, al igual que los
tipoterios, en cuyo suborden se les incluye a menudo, tenía formas
parecidas a los lagomorfos y a los roedores, que se convirtieron en
roedores eficaces. Los representantes más tardíos poseen un diastema
entre los incisivos y las muelas y todos los dientes crecían durante
toda la vida. Aparecieron poco después que los interatéridos, a
mediados del Eoceno, y no se extinguieron hasta el Plioceno,
alrededor de 3 millones de ańos atrás.
Interatherium excavatus.
Ameghino, 1887.
|
 |
 |
 |
|
Esqueleto de
Interatherium excavatus en el Museo Field de Historia
Natural, Chicago |
Cráneo y
mandíbula de
Interatherium robustum. Museo de
Paleontología de la Universidad de Córdoba, Argentina. |
Recreación de Interatherium excavatus
por Velizar Simeonowski. |
Mamífero Notoungulado.
Es un género extinto de mamíferos notoungulados de
la familia Interatheriidae. Vivió a principios del Mioceno en lo que
hoy es América del sur. La mayoría de los interaterios eran
mamíferos bastante pequeńos, parecidos a los roedores. Fueron un
grupo longevo, con representantes fósiles desde fines del Paleoceno
hasta fines del Mioceno. Numerosos materiales de la familia fueron
recolectados de 12 niveles estratigráficos, situados dentro de los
120 metros inferiores de la Formación Santa Cruz en la zona de la
costa atlántica, entre los ríos Coyle y Gallegos, en Estancia La
Costa, Cańadón Silva, Corriguen Aike (Puesto de la Ea. La Costa) y
Monte Tigre (Estancia Angelina). Otro género bien conocido es Interatherium,
del cual el resto fósil más abundante pertenece a Interatherium
robustum.
Colpodon
propinquus. Burmeister 1885.
|
 |
|
 |
|
Cráneo de
Colpodon, con detalles de su maxilar. Colección del MEF.
Procedente del Mioceno de Chubut. |
|
Posible
aspecto de Colpodon. Por MarMag.2026 |
Mamífero Notoungulado.
Es un género extinto de mamífero de la clase de los
notoungulados que habitaron América del Sur durante casi la mitad
del periodo Mioceno, hace 25 millones de ańos, perteneció a la
subfamilia de los leontiniidae el cual se extinguió casi al terminar
dicho periodo. Vivió en llanuras con alta vegetación y donde pasaban
gran parte del tiempo pastando. Su tamańo y dentición recuerdan a
los actuales rinocerontes. Eran animales con una estructura corporal
poderosa. Eran cuadrúpedos. La familia se identifica por poseer
incisivos parecidos a los de los cánidos. El resto de la dentadura
suele ser braquiodonte. Este animal es conocido casi exclusivamente
por restos craneales y, por lo tanto, la apariencia general es
bastante hipotética. En comparación con animales similares más
conocidos (por ejemplo, Scarrittia y Huilatherium),
se cree que Colpodon era un animal de formas
relativamente pesadas, con una estatura similar a la de un carnero.
El cráneo era bastante corto y alto, dotado de una dentición inusual
en comparación con el de la mayoría de sus parientes cercanos (los
leontínidos). Colpodon es uno de los primeros
notoungulados conocidos; fue descrito por primera vez en 1895 por
Burmeister; el erudito basó el género Colpodon en un
molar superior y dos molares inferiores encontrados en la
desembocadura del río Chubut (Argentina). Burmeister consideró Colpodon una
forma intermedia entre Nesodon y Homalodotherium.
Más tarde el propio Burmeister describió y representó otros restos
de la especie tipo (Colpodon propinquus). A principios
de la década de 1900, Florentino Ameghino describió otras especies
de Colpodon, como Colpodon
distanceus y Colpodon plicatus.
Este último creyó que este animal era un representante de los
leontínidos.
|
Diadiaphorus majusculus. Ameghino, 1887.
|
|
Diadiaphorus
Diadiaphorus Diadiaphorus
Diadiaphorus Diadiaphorus
Diadiaphorus Diadiaphorus
Diadiaphorus |
|
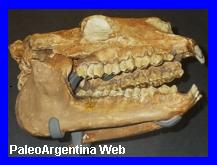
Cráneo y
mandíbula de Diadiaphorus del Mioceno de Chubut. Al lado,
detalle de la estructura de la extremidad.
(*) |
 |

Aspecto de
equino que presentaba el extinto Diadiaphorus, un
notoungulado extinto de Sudamérica.
(*) |
Mamífero
Litopterno.
La difusión de las planicies abiertas por todo el
continente sudamericano contribuyo a la evolución de unos animales
corredores de estructura ligera. Los proteroterios (primeras
criaturas) eran animales con aspecto de caballos, que vivieron entre
el final del Paleoceno hasta el final del Plioceno, e incluso, en el
Pleistoceno medio según nuevas evidencias. Estos han experimentado
cambios evolutivos y adaptaciones paralelos a los verdaderos
caballos de Norteamérica, incluso mucho antes que en cualquier otro
sitio. El Diadiaphorus debió tener un aspecto muy
semejante al antílope de cuello corto o un poney. Tenía el tamańo
semejante al de una oveja, pero con las patas de un caballo de tres
dedos. Aunque los huesos pares de las extremidades (cubito, radio y
tibia, peroné) nunca llegaron a fundirse, como ocurrió después con
los caballos auténticos, las patas eran largas y delgadas. El digito
central (el tercero) era muy grande y soportaba todo el peso del
animal, mientras que los dos situados en cada lado (el segundo y el
cuarto) se habían atrofiado. La cabeza era corta y gruesa, y la caja
craneal era bastante grande. Debido a que las coronas dentarias eran
bajas, es probable que Diadiaphorus ramoneara la
vegetación más blanda, como arbustos y los árboles de las planicies
patagónicas. Otra especie conocida; Diadiaphorus caniadensis (Kramarz
& Bond, 2005).
Theosodon garretorum.
Scott, 1910.
|
 |
 |
 |
|
Cráneo y mandíbula de
Thesodon. MNHN. (*)
|
Mandíbula de Theosodon.
Museo de Paleontología de la Univ. de Córdoba. |
Reconstrucción en vivo
de Thesodon. (*)
|
Mamífero
Litopterno.
El género vivía en lugares abiertos, como los actuales
guanacos, al cual se asemejan. Poseían un cuello largo y una trompa como
la de los tapires, aunque no guarda relación con ninguno de ellos. Su
parecido es resultado de convergencia adaptativa. Los teosodontes, como
la mayoría de los meridiungulados, eran exclusivos de América del Sur.
Trotando por las llanuras, estas criaturas de cuello largo, que
ramoneaban y también pacían, debieron parecerse mucho a las manadas de
Auquénidos actuales. Exteriormente, la diferencia principal debía estar
en las patas. Las especies del género Theosodon tenían
tres dedos y eran, por ende, más pesadas. La posición retraída de las
fosas nasales en el cráneo, indica que también tenían trompa, aunque tal
vez no más prominente que la del antílope saiga actual. La mandíbula
inferior era muy delgada y en la boca, como en el resto de los
litopternos, había una dentadura completa, formada por 44 piezas, el
máximo para los mamíferos placentarios, lo cual es poco frecuente en los
cuadrúpedos herbívoros correspondientes a una etapa tan tardía del
Terciario. De huesos finos, las especies variaban aproximadamente de 1 a
2 metros de altura y se estima que pesaban desde cerca de 40 kilogramos
en las especies más pequeńas, hasta 140 kilogramos en las mayores.
Probablemente vivieron en las planicies, en grupos, alimentándose de
gramíneas y hojas de árboles y arbustos. Existieron durante el Mioceno y
el Plioceno, hace entre 13 a 5 millones de ańos. El género de los
teosodontes es un representante de la familia de los Macraucheniidae,
junto a varios otros como Scalabrinitherium, del cual
evolucionó el más conocido entre los géneros de la familia, Macrauchenia del
Pleistoceno, descrito por Darwin. La gran mayoría de especies parecen
haber desaparecido durante el Plioceno coincidiendo con el Gran
Intercambio Biótico Americano.
Brachytherium cuspidatum. Ameghino 1883.
Nomen dubium
|
 |
|
 |
|
Fragmento mandibular de
Brachytherium,
del Mioceno superior de la Prov. de Buenos Aires. Museo de
La Plata. |
|
Aspecto de
Brachytherium,
una forma de evolución paralela con los equinos. (*). |
Mamífero
Litopterno. Animales
de pequeńo y mediano tamańo que por su aspecto y sus proporciones
eran caballos en miniatura, como ocurrió después paralelamente entre
los équidos arcaicos. Los primeros proteroterios tenían un dedo
central muy grande en cada pie, y dos laterales más pequeńos que
apenas tocaban al suelo, algo similar a lo que ocurre en las patas
traseras de los jabalíes actuales. En especies más recientes estos
dedos laterales fueron reduciéndose cada vez más. Se piensa que
evolucionaron de litopternos procedentes de la familia
Notonychopidae, cuya familia vivio entre el Oligoceno y el final del
Pleistoceno. Hubo otra familia de meridiungulados, perteneciente al
suborden de los Notoungulata que también presentaron formas
semejantes a los caballos. La difusión de las planicies abiertas por
todo el continente sudamericano contribuyó a la evolución de unos
animales corredores de estructura ligera. Parece que los
proteroterios hubieron de experimentar buena parte de los mismos
cambios adaptativos que los caballos primitivos de América del
Norte, algunas veces antes que los desarrollos que se producían en
cualquier otro sitio. Es poco probable que fueran capaces de
pastear, sin embargo, ya que su dentadura parece corresponde a
animales ramoneadores, alimentándose de arbustos y, adaptados a la
carrera. Son muy similares a la de la subfamilia extinta de caballos
Anchitheriinae, que fue abundante en el Mioceno de Eurasia.
Oxyodontherium zeballozi.
Ameghino 1883.
|
 |
|
 |
| Cráneo de
Oxyodontherium zeballosi, exhibido en el Museo de Cs
Naturales y Antropológicas de Mendoza. |
|
Aspecto de un
macrauquénido (*) |
Mamífero
Litopterno.
Era un animal más robusto y corpulento, como un
caballo, de miembros largos, con tres dedos en cada pie, como los
tapires y un cuello largo como el de una jirafa, cuyo nicho
ecológico ocupaba con formas intermedias que recuerdan al okapi, con
un cráneo pequeńo y provisto, en vida, de una corta trompa parecida
a la de un tapir. Estos macrauquénidos son una familia extinta de
mamíferos placentarios del orden Litopterna perteneciente al
superorden Meridiungulata. La dentadura de todo el grupo presenta la
dotación de los primeros mamíferos, con 44 dientes. El nicho
ecológico fue posteriormente ocupado por litopternos de mayor
tamańo, todos extintos, y en la actualidad en parte por el guanaco,
que ocupo su nicho ecologico. Otras especies relacionadas; Macrauchenia
minuta (Burmeister, 1885), Mesorhinus piramydatus (Ameghino, 1885)
y Oxyodontherium piramydatus (Ameghino, 1885).
Huayqueriana cristata. Rovereto,
1914.
|
 |
 |
|
Vista ventral del cráneo de
Huayqueriana del Mioceno de Mendoza. |
Huayqueriana, según el
paleoartista Jorge Blanco. |
Mamífero
Litopterno. Las
diversas familias de ungulados nativos de América del Sur
prosperaron los Litopterna desde fines del Paleógeno al comienzo del
Holoceno. La Formación Huayquerías (Mioceno tardío) está ampliamente
expuesta en el centro de Mendoza. En expediciones paleontológicas en
la primera mitad del siglo XX, han dado recientemente un número
significativo de nuevos hallazgos fósiles. Los macrauquenios son, en
su mayor parte, animales semejantes a los jiráfidos o a los
camellos. Sus dientes son, por lo general, más sencillos que los de
los ungulados de otros lugares; la dentadura seguía siendo completa
y el intersticio o diastema entre los dientes anteriores y las
muelas no estuvo nunca tan desarrollado. Las extremidades presentan
siempre tres dedos como en los rinocerontes o ceratomorfos,
pertenecientes al orden de los perisodáctilos, con los que no están
relacionados evolutivamente. Uno de los aspectos más llamativos del
cráneo de los macrauchénidos son las grandes aperturas nasales
ubicadas en el dorso del hocico y vertical al paladar. La excelente
preservación del ejemplar del IANIGLA-Mendoza, permitió, con la
ayuda de tomografías computadas de alta resolución, estudiar en
detalle la anatomía interna, incluso explorar su cavidad nasal.
Proterotherium
cervioides.
Ameghino, F. 1883.
|
 |
 |
 |
|
Mandíbula de Proterotherium cavun.
Museo de Paleontología de la
Universidad de Córdoba. |
Molde
endocraneano natural de Proterotherium cavum(*). |
Un ave de
terror persigue a un grupo de Proterotherium. Por
MarMag.2026 |
Mamífero
Litopterno.
Es un género de litopterno, mamíferos
ungulados extintos de la familia de los proterotéridos que
existieron durante el Mioceno. Tuvo una amplia distribución en
Sudamérica, hallándose sus fósiles en Argentina, Chile y Brasil.
Animales de pequeńo y mediano tamańo que por su aspecto y sus
proporciones eran caballos en miniatura, como ocurrió después
paralelamente entre los équidos arcaicos. En especies más recientes
estos dedos laterales fueron reduciéndose cada vez más. Los
proteroterios se piensa que evolucionaron de litopternos procedentes
de la familia Notonychopidae. La difusión de las planicies abiertas
por todo el continente sudamericano contribuyó a la evolución de
unos animales corredores de estructura ligera. Parece que los
proteroterios hubieron de experimentar buena parte de los mismos
cambios adaptativos que los caballos primitivos de América del
Norte, algunas veces antes que los desarrollos que se producían en
cualquier otro sitio.
|
 |
|
 |
|
Mandíbula inferior de
Licaphrium floweri.
Museo de
Paleontología de la Universidad de Córdoba. |
|
Aspecto de
Licaphrium floweri.
(*). |
Mamífero
Litopterno.
La enorme extinción de llanuras
abiertas por todo el continente sudamericano contribuyó a la
evolución de unos animales corredores de estructura ligera. Como los
anteriores, eran animales de pequeńo y mediano tamańo que, por su
aspecto y sus proporciones, eran caballos en miniatura, como ocurrió
después paralelamente entre los équidos arcaicos. Los primeros
proteroterios tenían un dedo central muy grande en cada pie, y dos
laterales. Algunos fósiles bien preservados de Licaphrium, proceden
de Puesto Estancia La Costa. Un sinónimo también es conocido como Anisolophus
fischeri Mercerat 1891. Otras especies conocidas son; Licaphrium
debile, Licaphrium floweri, Licaphrium granatum, Licaphrium
intermedium, Licaphrium intermissum y Licaphrium
mesopotamiense.
|
Homunculus patagonicus. Ameghino,
1894.
|
|
Homunculus patagonicus Homunculus patagonicus Homunculus
patagonicus Homunculus patagonicus Homunculus
patagonicus
|
|

|

|

|
|
Cráneo del
primate Homunculus in situ en el sedimento.
(*) |
Cráneo de
Homunculus patagonicus depositado en el Museo de
Paleontología de Córdoba.
|
Reconstrucción
en vivo. Por MarMag.2026 |
Mamífero Primate.
Es la única especie del género
monotípico extinto de primate platirrino Homunculus,
el cual vivió en la Patagonia argentina, en el sur de América del
Sur, durante el Mioceno inferior. Fue un primitivo primate (monos)
aloctono, cuyo ingreso al continente fue hace 30 millones de ańos
según la evidencia fósil reunida hasta el momento. Vivió en lo que
es hoy el Rió Pinturas, en la Patagonia Argentina durante la edad
Santacrucense. Los hallazgos de estos mamíferos indican que durante
parte del Mioceno temprano, el sur argentino tenia condiciones
ambientales muy semejantes a las que podemos observar en la selva
misionera y amazónica. Su tamańo es muy pequeńo y no pesarían más de
1 kilogramo. Al igual que sus representantes vivientes, su dieta era
omnívora. Comían insectos, frutos, pequeńos mamíferos etc. Lo que
más llama la atención de estos primates es su "encefalizacion
progresiva", es decir, el aumento de la corteza cerebral a
comparación de su tamańo, lo que le permitiría retener información e
inteligencia, pero en este periodo algo muy remoto. La especie tenía
el rostro relativamente corto y órbitas de moderado tamańo, lo cual
sugiere que se trataba de una especie con hábitos diurnos. El cráneo
era grácil y carecía de cresta sagital. Tenía una masa corporal de
alrededor de 2,7 kilogramos. Basándose en la morfología dental, se
trataba de una especie frugívora que complementaba con hojas para
suplir su dieta. De acuerdo a los restos postcraneales,
probablemente se trataba de un cuadrúpedo. También se lo conoce con
el sinónimo Anthropops perfectus. Otros primates del
Mioceno de Argentina son; Dolichocebus gaimanensis Kraglievich,
1951, Soriacebus ameghinorum Fleagle et al., 1987, Soriacebus
adrianae Fleagle, 1990; Carlocebus carmenensis Fleagle,
1990; Carlocebus intermedius Fleagle, 1990; Proteropithecia
neuquenensis Kay et al., 1998, Mazzonicebus almendrae Kay.
2010; y Tremacebus harringtoni Rusconi, 1933
|
Killikaike blakei.
Tejedor, 2006. |
|
Killikaike blakei Killikaike
blakei Killikaike blakei Killikaike blakei Killikaike blakei
Killikaike blakei Killikaike blakei Killikaike |
|
Material
in-situ del primate. El fósil del cráneo de
Killikaike y reconstrucción del aspecto
original.
(*). |
Mamífero Primate.
Un grupo de
paleontólogos ha hallado en la Patagonia Argentina un cráneo de un
primate que aporta relevantes datos sobre la evolución del tamańo
cerebral de esta especie. Los resultados del estudio del que está
considerado el mejor resto craneal de un primate preservado en
Argentina aparecieron publicados en la revista PNAS, firmados por el
equipo que dirige el paleontólogo Tejedor en la Facultad de Ciencias
Naturales de la Universidad Nacional de la Patagonia "San Juan Bosco".
Los primates sudamericanos nada tienen que ver con el origen del
hombre, se encuentran ampliamente distribuidos y son diversos en
Centroamérica y Sudamérica, pero su registro fósil es sumamente
escaso, aparte de que los restos se fragmentan con facilidad. El
nuevo cráneo descubierto muestra un rostro completo y sin
deformaciones; el hueso frontal conserva intacto el espacio interior
de los lóbulos frontales del cerebro y se mantiene además la mayor
parte de los dientes superiores casi sin desgaste. El fósil
corresponde a un nuevo género y especie de primate denominado Killikaike
blakei, en homenaje a la estancia de procedencia (Killik
Aike Norte) y a sus propietarios, los seńores Blake. Lo más
sorprendente detectado en el primate Killikaike blakei son
las avanzadas y modernas proporciones de su cerebro, unos rasgos que
desvelan que su gran crecimiento cerebral habría ocurrido
aproximadamente doce millones de ańos antes que en los ancestros
africanos del hombre. No obstante, el rostro y los dientes aún
presentan ciertos caracteres primitivos, lo que podría implicar que
sus habilidades cognitivas probablemente precedieron a las
relacionadas con hábitos alimentarios. El primate fue localizado
cubierto por una roca de ceniza volcánica y sólo se podía apreciar
una mínima parte de la superficie dental. El primer paso fue
verificar, mediante una tomografía computerizada, si dentro de la
roca había realmente un cráneo y, el segundo, corroborar el lugar de
procedencia. Esta segunda tarea fue sencilla, porque en las
cercanías había un yacimiento con fósiles procedentes precisamente
de un nivel de cenizas volcánicas, donde el pasado ańo se halló un
espécimen adicional de este género. Ello permitió un análisis
químico de la roca ubicada alrededor del cráneo y se comprobó que la
misma tenía una antigüedad de hace 16.4 millones de ańos,
coincidente con el Mioceno temprano, período del Terciario que
comenzó hace 23 millones de ańos y concluyó hace unos 6 millones de
ańos. Además, se concluyó que el nuevo fósil de primate era de la
subfamilia de los cebinos, en la que se incluyen los actuales monos
ardillas (género Saimiri) y los capuchinos (género Cebus).
La particularidad de los cebinos es su cerebro muy desarrollado
respecto al tamańo corporal, lo que también se da en los ancestros
humanos, los homínidos africanos. Los monos ardillas y capuchinos
manifiestan comportamientos asociados con una notable inteligencia,
viven en complejos grupos sociales e, incluso, los capuchinos pueden
manipular instrumentos rudimentarios.
Stegotherium tessellatum.
Ameghino,
1887.
| |
|
|
|
 |
 |
 |
|
Stegotherium tessellatum, del Mioceno
de la formación Santa Cruz. Princeton University.
|
Coraza dorsal
y cefálica de Stegotherium,
en el Geoparque Bryn Gwyn de
Gaiman, Chubut. |
Aspecto de
Stegotherium tessellatum.
Por MarMag.2026 |
Mamífero
Xenarthro.
Es un género extinto de armadillos de
gran tamańo de la familia Dasypodidae. En el ańo 1887, Florentino
Ameghino describió de manera preliminar el primer armadillo
Stegotheriini, Stegotherium tessellatum, a base de
placas de la coraza. Se trata de un armadillo primitivo que vivió
en la Patagonia argentina entre el Eoceno al Mioceno inferior, el
cual presenta un cráneo alargado y prolongado, con forma de pico y
mandíbulas esteliformes como los oso hormigueros, pero con algunos
dientes rudimentarios. Su coraza estaba formada por placas óseas
sueltas, no soldadas, presentando el sistema pilífero sumamente
desarrollado. Los dasipodidos se caracterizan por poseer un
caparazón dorsal formado por placas yuxtapuestas, ordenadas por lo
general en filas transversales, con cola bastante larga y
extremidades cortas. Los dientes del armadillo son estructuras
cilíndricas simples y uniformes, sin esmalte y con raíces abiertas y
crecimiento continuo. Los armadillos Stegotheriini se encuentran
representados en unidades Cenozoicas desde el Eoceno de la Patagonia,
(Edad Casamayorense) hasta el Mioceno medio de Argentina y Chile.
|
 |
 |
 |
|
Coraza de
Proeutatus, en el
Geoparque Bryn Gwyn de
Gaiman, Chubut. |
Fragmento de
cráneo de Proeutatus oenophorum.
Museo de Paleontología de la Universidad de
Córdoba. |
Aspecto de
Proeutatus. Por MarMag.2026 |
Mamífero
Xenarthro.
Fue un dasipodido muy común en el Mioceno de
Patagonia, que recuerda en cierta forma con el Tatu Carreta. Al
igual que los armadillos, pose un escudete de placas óseas en su
cráneo, conocido también como escudete cefálico. Su coraza dorsal es
robusta, y poseía unas 33 bandas móviles que cubrían las dos
terceras partes del caparazón. En cada mandíbula poseía de 9 a 10
dientes, los cuales utilizaba para comer carne en descomposición,
huevos, larvas, caracoles y algunos tallos. Es muy posible que Proeutatus
oenophorum, se alimentara de pastizales de llanuras abiertas de la
antigua Patagonia. Construían grandes galerías subterráneas, con
cámaras amplias para proteger sus crías. Su cráneo era alargado. Sus
patas eran cortas y robustas, protegidas por unas enormes falanges
ungueales o garras, las cuales, eran justamente utilizadas para
cavar.
Cochlops muricatus.
Ameghino,1889.
|
Craneo y coraza.
Material colectado en 1896 en la provincia de Santa Cruz y
depositados desde entonces en
Yale Peabody Museum of Natural History.
Ilustración de Marcelo Canevari. |
Mamífero
Xenarthro. La
familia gliptodontes constituye uno de los grupos de mamíferos
fósiles más comunes y diversificados del Cenozoico sudamericano, y
está representada por aproximadamente 65 géneros. Los primeros
representantes de esta familia se extienden desde el Eoceno tardío
al Holoceno temprano. Cochlops, vivió en el Mioceno
medio-tardío (hace aproximadamente 18 a 12 millones de ańos) y sus
restos fósiles fueron encontrados en América del Sur. Este animal,
como todos los gliptodontes, estaba equipado con una coraza formada
por numerosos osteodermos soldados entre sí, que protegían la mayor
parte del cuerpo. El cráneo de Cochlops se caracterizó
por un rostro más corto, con el perfil facial fuertemente inclinado
hacia el frente y desde el plano occipital oblicuo. En general, la
forma del cráneo parece anunciar la de gliptodontes posteriores,
como el de Panochthus. El caparazón de Cochlops se
caracterizó por placas particularmente arrugadas, especialmente en
comparación con otras formas de gliptodontes arcaicos como Asterostemma o Propalaehoplophorus.
Algunas placas o osteodermos, especialmente en la región pélvica,
mostraron una ornamentación particular, con un gran tubérculo cónico
central rodeado de tubérculos arrugados más pequeńos. El género
Cochlops fue descrito por primera vez por Florentino Ameghino en
1889, sobre la base de restos fósiles encontrados en Argentina, en
suelos del Mioceno inferior. La especie es Cochlops muricatus,
pero el género Cochlops debilis, descrito por el
propio Ameghino en 1891, también se atribuyó a este género.
Glyptatelus tatusinus.
Ameghino, 1897.
|
 |
|
 |
|
Fragmentos de placas óseas
de Glyptatelus. (*) |
|
Aspecto en vida de
Glyptatelus por Julio Lacerda. (*). |
Mamífero
Xenarthro.
Pertenecen a una familia muy particular de
Xenarthros extinguidos, los "Glyptodontidae". Tenían una coraza
ósea (Cingulata) autóctonos de América del sur, de tamańo
mediano, alcanzando una notable diversificación durante el
Plioceno y Pleistoceno. Su caparazón óseo, espeso y rígido,
formado por placas soldadas entre sí, con dentición
especializada para la dieta herbívora, alimentándose
principalmente del pastoreo. Se convirtieron rápidamente en
animales exitosos para los pronunciados cambios climáticos que
se aproximaban. Su coraza no les permitía realizar grandes
movimientos ni realizar cuevas, ya que estas no presentaban
bandas móviles como los peludos y mulitas actuales. Su cola
probablemente estaría provista de unos anillos óseos, como los
ejemplares más recientes, tanto extintos como vivientes, pero la
ausencia por ahora de estos restos no afirma que sea así. Su
cráneo era grande y tosco, cubierto en la parte superior por un
escudete de placas, lo que protegía al animal cuando era atacado
por predadores de la época, como las gigantescas aves o los
marsupiales dientes de sable. Sus patas eran muy cortas y
robustas como para soportar el peso del animal. Su fémur
totalmente desproporcionado a comparación a otros huesos largos,
y la pelvis soldada al caparazón del animal. El mismo vivió
durante el Mioceno de Chubut, Argentina.
Phlyctaenopyga ameghini.
Ameghino, 1889.
|
 |
 |
 |
| Coraza de
Phlyctaenopyga ameghini, precedente
del Mioceno tardío del Noroeste de Argentina. Museo de La
Plata. |
Coraza y
otros restos de una nueva especie de
Phlyctaenopyga.
Museo Rincón de Atacama
de Termas de Rió Hondo. |
Aspecto del gliptodonte Phlyctaenopyga.
Imagen tomada de E loy
Manzanero. |
Mamífero
Xenarthro.
Como todos los Gliptodontes, este animal también
tenía un caparazón robusto hecho de placas óseas fusionadas,
cubriendo la mayor parte del cuerpo. Phlyctaenopyga era
un gliptodonte de tamańo mediano, y generalmente podía superar los
dos metros de longitud. El caparazón consistía en osteodermos con
una figura central rodeada por dos o tres filas de figuras
periféricas; la segunda fila estaba completa solo en algunos
osteodermos, mientras que la tercera fila siempre estaba incompleta.
Además, el cráneo era muy corto y ancho, especialmente en la región
del hocico. Las placas que protegían la región de la cabeza estaban
prácticamente desprovistas de ornamentación. En el Mioceno tardío de
Catamarca, se registraron importantes restos de Phlyctaenopyga
ameghini. Recientemente se dio a conocer una nueva especie
de Phlyctaenopyga proveniente de la Formación Las
Cańas (Mio-Plioceno) de las Termas de Río Hondo, en la Provincia de
Santiago del Estero. El material consiste en una coraza dorsal casi
completa, dos anillos caudales y un fragmento de la región media del
tubo caudal, alojado en el Museo Municipal Rincón de Atacama de la
localidad termense. Al igual que en otros gliptodontes, fue un gran
herbívoro. Otras especies relacionadas; Phlyctaenopyga trouessarti, Nopachtus
coagmentatus, Stromaphorus y Pseudoplohophorus.
Cranithlastus xibiensis.
Arias et al. 1978
|
 |
|
 |
|
Ejemplar tipo de Cranithlastus xibiensis, en el Museo
de Ciencias Naturales de la Universidad de Salta. |
|
Posible aspecto de Cranithlastus xibiensis, similar a
otros géneros mas tardíos.
Ilustración de
Carlos Alberto Montefusco. |
Mamífero
Xenarthro.
Es una especie de mamífero acorazado, conocido
popularmente como Gliptodonte. El primer ejemplar conocido, está
conformado por un esqueleto parcial. Su localidad tipo es Barrancas
del Río Chico, que se encuentra en una arenisca arcillita,
huayqueria en la Formación Piquete de la provincia de Jujuy,
Argentina, cuyos restos fueron preparados y preservados en el Museo
de Ciencias Naturales de la Universidad de Salta. Posteriormente, en
2017, se dieron a conocer nuevos restos de Cranithlastus
xibiensis, del Mioceno Tardío en el río Xibi Xibi, Jujuy.
Los restos de fósiles recuperados, corresponden a la parte anterior
de una coraza dorsal, escudete cefálico y el cráneo mayormente
completo, correspondientes a un único individuo. Su cola estaba
formada por anillos móviles y un tubo caudal más cilíndrico que en
otros géneros. Este como otros gliptodontes, conforman la conocida
fauna de mamíferos acorazados de Sudamérica.
Pelecyodon cristatus.
F. Ameghino,
1891.
|
 |
 |

|
|
Cráneo y
mandíbula de Pelecyodon cristatus, hallado en el extremo
austral de Patagonia, Estrecho de Magrallanes.
Museo de Paleontología de la Universidad de Córdoba. |
Cráneo completo y mandíbula inferior
de
Pelecyodon del Mioceno
temprano de Santa Cruz. Museo de La Plata. |
Aspecto en vida
de Pelecyodon cuando la Patagonia era una selva
tropical. (*) |
Mamífero
Xenarthro.
Durante el Mioceno,
hace 20 millones de ańos, los Xenarthros estaban representados y
adaptados en América del sur, el cual, en el Mioceno se encontraba
geográficamente bien distribuido por la actual Patagonia Argentina. Pelecyodon era
de tamańo chico, media hasta un metro de largo, a diferencia de sus
gigantescos parientes del Pleistoceno que superaban los cuatro
metros de altura. Tenía garras muy desarrolladas y curvas (una
característica común entre los mamíferos edentados) seguro debe
haberse visto desplazarse sobre sus nudillos anteriores. Es muy
probable que pasara la mayor parte de su tiempo en las copas de los
árboles, sujetándose de sus garras y alimentándose de frutos
suculentos en los tupidos bosques sub tropicales del Mioceno. Como
todos los edentados, Pelecyodon tenía pocos dientes y
sin esmalte. Sus mandíbulas solo tenían cuatro pares de dientes. Si
bien hoy en día la Patagonia Argentina es un gran desierto, durante
el Mioceno estaba constituida por grandes y extensas selvas,
evidenciado por la diversidad faunistica y botánica recuperada. Este
animal está emparentado con el Perezoso de Tres Dedos (Bradypus),
que vive actualmente desde el norte de Argentina a Nicaragua. Otros
géneros relacionados; Eucholoeops ingens y Pseudhapalops s.p.
Eucholoeps
fronto. Ameghino, 1887.
|
 |
 |
 |
|
Cráneo y mandíbula (frontal) de
Eucholoeps fronto.
Museo de Paleontología de la
Univ de Córdoba. |
Cráneo completo de
Eucholoeps ingens
del
Mioceno temprano de Santa Cruz. Exhibido en el Museo de La
Plata. |
Aspecto del
perezoso arborícola del Mioceno patagonico.
Eucholoeps .
Por MarMag.2026. |
Mamífero
Xenarthro. Los
megaloníquidos son una familia de mamíferos placentarios del orden
Pilosa. Fue muy diversa durante el Cenozoico, pero en la actualidad
solo sobrevive un género, Choloepus, con dos especies.
Aparte de varios restos óseos, también se conoce un encéfalo de Eucholoeps
fronto, se interpreta en base a un molde endocraneano del
Mioceno temprano, procedente de Monte León Santa Cruz. Era un
perezoso tridáctilo. Poseía pelo más largo seguramente, ojos más
grandes, y sus patas traseras y delanteras son menos
desproporcionadas en tamańo. Su cola, sin embargo, es más corta. Su
dentición es relativamente escasa. Se alimentaba de hojas tiernas y
seguramente era arborícola. La presencia de perezosos pequeńos y
monos indica que en la Patagonia existían frondosos bosques. Otra
especie conocida; Eucholoeps ingens.
|
|
|
|
Protamandua
Protamandua Protamandua Protamandua |
Protamandua
Protamandua Protamandua Protamandua |
|

|

|
|
Cráneo de Neotamandua sp
del Mioceno del Noroeste argentino. Museo de La Plata. |
Aspecto en vivo de Protamandua.
Por MarMag.2026 |
Mamífero
Xenarthro.
Genero de mamífero Xenarthro (Vermilingua) de tamańo
mediano a moderadamente grande, terrícolas y semiarbolicolas. Es muy
probable que, al igual que sus representantes actuales y sus
parientes gigantes del Pleistoceno, tuviera un pelaje formado por
pelos cerdosos, espesos y duros. Poseían una cola muy larga, la cual
duplicaba la longitud total. Su cráneo es muy particular y a veces
cuesta a simple vista identificarlo como tal. El mismo tiene forma
alargado, delgado, ligeramente curvado en la parte inferior y
termina en un hocico romo. Su maxilar tiene la misma longitud,
siendo muy poco movible el inferior, ya que su boca se vería como
una pequeńa hendidura. Según las características morfológicas del
cráneo podemos decir que sus ojos y orejas eran muy pequeńas. Su
columna vertebral es algo corta y ancha. Sus extremidades cortas.
Las delanteras tienen cuatro dedos provistos de unas falanges
úngeles muy robustas y comprimidas, las cuales se asemejan a las
garras de un ave rapaz. Las patas posteriores son menos robustas que
las anteriores, tienen pies con cinco dedos. Su lengua seria larga y
estrecha, la que serviría para alimentarse de hormigas, termitas y
otros pequeńos insectos, con la cual llegaría hasta los rincones
menos accesibles. Es probable que tuviera un metabolismo algo lento.
Los fósiles de este llamativo mamífero es algo escaso, sus restos
aparecen en el Mioceno del Noroeste, Plioceno de Patagonia y en la
Provincia de Buenos Aires.
Nematherium angulatum.
Ameghino, 1887.
 |
|
 |
|
Cráneo de
Nematherium sp en el Royal Ontario Museum. (*) |
|
Aspecto en
vida de Nematherium. (*) |
Mamífero
Xenarthro.
Los pilosos son un orden de mamíferos
placentarios que incluye los osos hormigueros, los tamandúas y los
perezosos. En la actualidad existen únicamente en el continente
americano. El origen del orden se remonta a principios del Terciario
(hace alrededor de 60 millones de ańos, poco tiempo después de la
extinción de los dinosaurios). El género Nematherium de
tamańo algo mayor a otros xenarthros, unos 2,5 metros de largo,
aunque la ausencia de fósiles, no ayuda a tener la suficiente
información. Serían de ambientes cerrados, consumidores de hojas,
frutos y posiblemente tubérculos o raíces gracias a su mayor
capacidad cavadora. Probablemente era de movimientos lentos, usando
sus garras delanteras como arma defensiva. Aunque se consideró
vegetariano, algunos científicos plantean la posibilidad de que
hubiese consumido alternativamente carrońa, tema aun en discusión.
Posiblemente al igual que otros perezosos, exhibe la interesante
característica de poseer pequeńos osteodermos, huesos insertos en la
piel, que presumiblemente ańadirían alguna protección al animal. Sus
restos fueron recuperados en Formación Santa Cruz, Mioceno temprano
de la Patagonia Argentina. Otras géneros relacionados; Analcitherium Ameghino
(Mylodontidae) y Prepotherium Ameghino (Megatheriidae).
|
Palaeomyrmidon incomtus.
Rovereto, 1914.
|
|
Palaeomyrmidon
Palaeomyrmidon Palaeomyrmidon Palaeomyrmidon Palaeomyrmidon
Palaeomyrmidon Palaeomyrmidon |
Mamífero
Xenarthro.
Es un género extinto de oso hormiguero. Su pariente
vivo más cercano es el oso hormiguero enano (Cyclopes
didactylus). Aunque el oso hormiguero enano es arborícola,
parece que Palaeomyrmidon era más terrestre. Palaeomyrmidon es
conocido a partir de un cráneo fósil de 93 milímetros hallado en
Argentina, en la Formación Araucano. Este cráneo es más alargado que
el de Cyclopes y recuerda superficialmente al de los
tamandúas. Descrito por Rovereto en 1914, cuya especie tipo es
conocida por un cráneo casi completo, de unos 10 centímetros de
largo, procedente de sedimentos del Mioceno tardío de
Argentina. Genero monotípico de Xenarthros de tamańo similar al de
una ardilla, pero con aspecto similar al de un oso hormiguero
actual. Palaeomyrmidon fue estrictamente arborícola,
insectívoro y melivoro. Las patas de este singular fósil son
parecidas a otras especies ya mencionadas, por tener las delanteras
provistas de cuatro dedos, de los cuales dos tienen enormes y
afiladas falanges ungueales o garras. Este pequeńísimo animal vivió
en las abundantes selvas sub-tropicales de la antigua Patagonia,
donde podía estar bien refugiado de los principales depredadores de
la época. El tamańo de las fosas oculares seńalan hábitos nocturnos.
Es probable que fuera algo lento y de aspecto pesado, pero podía
trepar los árboles con cierta facilidad con su larga cola prensil,
al igual que lo hace un simio. Como hemos adelantado su alimentaron
estaba constituida por numerosos insectos, como hormigas, termes,
abejas, avispas y de las larvas de todos estos. Se podría poner en
dos patas, formando un trípode con su cola mientras que con las
patas delanteras se lavaba el alimento a la boca. Sus restos
provienen principalmente del Mioceno superior de Chubut, Argentina.
Prolagostomus pusillus.
Ameghino 1887.
|
 |
 |
 |
|
Restos de vizcacha fósil.
Ilustrativo. Museo Paleontológico de San Pedro. |
Vista del cráneo.
Ilustrativo. Museo de
Ciencias Naturales de Miramar. |
Reconstrucción en vivo.
Por MarMag.2026 |
Mamífero Roedor. Se trata de un
género bien reconocido en el Mioceno Argentino. Es un roedor
caviomorfo de tamańo relativamente mediano-pequeńo. Eran
herbívoros estrictos, aunque algunos autores sostienen probables
hábitos carrońeros, ya que, en sus paleocuevas o madrigueras
antiguas, con frecuencia se hallan restos óseos pertenecientes a
otros animales de la época con marcas semejantes a sus dientes,
pero también se puede tratar de una actividad propia de los
roedores, de roer cosas duras para evitar el crecimiento
descontrolado de sus incisivos. Este animal se encuentra
representado por otros dos géneros de su familia, uno de ellos
el Lagostomopsis antiquus del Plioceno, y Lagostomus
maximus del Pleistoceno y Holoceno. Se trata de especies
aloctonas que ingresaron al continente en el Oligoceno superior
adaptándose efectivamente a los nuevos cambios ambientales. A
este género lo conocemos por sus abundantes restos fósiles, ya
que no tiene descendientes vivientes. Para el Mioceno también se
conoce Lagostomus telenkechanum.
Phoberomys burmeisteri.
Ameghino, 1886.
|
 |
 |
 |
|
Fragmento de rama mandibular de
Phoberomys en el Museo de La
Plata. |
Cráneo de
Phoberomys recreado en
cartón con interesantes detalles, realizado por Luciano
Reuil. |
Tamańos
comparativos con un humano y un carpincho, el roedor
viviente de mayor dimensión. Por MarMag.2026 |
Mamífero Roedor. Era
un roedor caviamorfo endémico y autoctono (?) de América del
sur, de tamańo grande. Generalmente cuando hablamos de roedores
nos imaginamos las ratas o lauchas. Pero, Phoberomys burmeisteri era
algo más grande que un cerdo doméstico y superaría los 700 kilos
de peso, y 1,6 metros de altura a la cruz. Está emparentado con el súper roedor del Plioceno "Telicomys".
Su cuerpo era grande y de aspecto rechoncho. Sus extremidades
eran largas y angostas. Su maxilares y ramas mandibulares
estaban provistas por cuatro molares e incisivos con 2
centímetros de espesor.
Las coronas de sus molariformes nos hablan de un pastador/ramoneador.
Su dieta estaría adaptada a los follajes
más duros de la región. Estos ejemplares son solo conocidos por
piezas aisladas que no aportan mucha información sobre su
morfología. Sus principales restos proceden del Mioceno de Entre
Rios.
Eocardia
perforata.
Ameghino, 1887
 |
 |
 |
|
Fragmento mandibular de Eocardia s.p del Mioceno
temprano de Santa Cruz. Museo de La Plata. |
Cráneo y
mandíbula de Eocardia perforata.
Museo de Paleontología de la Universidad de
Córdoba. |
Posible aspecto de Eocardia s.p (*). |
Mamífero Roedor. Es un
género extinto de los roedores de América del Sur. Tenía
unos 30 centímetros de largo, y estaba relacionada con los
conejillos de Indias, cuises y el carpincho (el roedor viviente de
mayor tamańo). Su cabeza era grande y rectangular, así como cola muy
pequeńa o ausente. Poseían cuatro dedos en las extremidades
anteriores y tres en las posteriores. Los Eocardiidae son una
familia extinta de roedores histricomorfos que vivieron en
Sudamérica desde el Oligoceno superior hasta el Mioceno medio,
siendo Eocardia, el generó más representativo para
esta época. Parecen ser un taxón polifilético de Cavioidea basal. El
primer hallazgo de Eocardia fue realizado por
Florentino Ameghino por medio de un molar, el cual ubicó dentro de
la familia Toxodontidae. Ese mismo ańo, Ameghino reconoció el error
y lo coloco dentro de familia Caviidae, desde entonces sufrió varios
cambios taxonómicos hasta llegar a los ya mencionados. Este
animalito vivía entre los pastizales y refugios rocosos,
alimentándose de vegetales en zonas abiertas. Los roedores, habrían
viajado hasta Sudamérica en un momento climático óptimo, a mediados
del Eoceno. Por entonces, según creen otros investigadores, un
roedor africano habría logrado cruzar el océano en una balsa
vegetal y habría llegado hasta el nordeste de Brasil en una o dos
semanas, ya que, el mar que separaba ambos continentes era menos
extenso. Otros géneros relacionados: Asteromys, Chubutomys,
Eocardia, Luantus, Matiamys, Neophanomys, Phanomys y Schistomys.
Neoreomys australis.
Ameghino
1887.
|
 |
 |
 |
| Cráneo y
mandíbula de Neoreomys (*). |
Vértebras de
Neoreomys (*): |
Aspecto del roedor mioceno (*). |
Mamífero
Roedor. Es un género
de roedores histricomorfos de la familia Dasyproctidae. Seguramente
su hábitat era principalmente el sotobosque de las selvas tropicales
y los bosques de galería, que existían incluso en el actual desierto
patagónico durante el Mioceno. Se alimentaban de frutos y semillas,
especialmente de palmas, también de tubérculos y brotes de hierba.
Acostumbraba a enterrar semillas cada vez que se alimenta, para
comerlas cuando había escasez. Excavaban madrigueras con varias
galerías de más de 60 centimetros de extensión. Una importante
cantidad de restos y colecciones provienen principalmente de las
formaciones geológicas de Pinturas y La Cańada en la Provincia de
Santa Cruz. Otra especie conocida es Neoreomys pinturensis,
Neoreomys decisus, Neoreomys indivisus, Neoreomys leptorhynchus y Neoreomys
pachyrhynchus.
Steiromys detentus.
Ameghino,
1887.
|
 |
|
 |
| Fragmento de
rama mandibular de
Steiromys detentus.
Museo de Paleontología de la Universidad de
Córdoba. |
|
Aspecto de
Steiromys detentus.
Por MarMag.2026 |
Mamífero Roedor. Es
un género de roedores histricomorfos de la familia Erethizontidae.
Incluye a una serie de especies conocidas con el nombre de coendús,
puercoespines americanos de cola larga ampliamente distribuidos por
la región Neotropical (América Central y del Sur). La característica
principal es su espalda cubierta de espinas que les sirve como
mecanismo de defensa, ya que cuando se sienten amenazados, las
erizan y apuntan hacia sus enemigos. En la actualidad, son de
hábitos nocturnos, vive en las ramas de los árboles de las que casi
nunca bajan, solitario o en pareja. De día duerme entre los árboles
huecos. Prefiere los niveles altos y medios del bosque con lianas.
No salta, y para cruzar un espacio entre dos árboles, debe descender
hasta el suelo. Se alimenta de hojas, frutos y ramas verdes y
pequeńas. Sus representantes fósiles como Steiromys detentus seguramente
tendrían un comportamiento similar. Algunos restos proceden de
la Estancia Ana María, Formación Pinturas, Lago Buenos Aires, en la
provincia patagónica de Santa Cruz. En los mismos estratos se han
recuperado restos de perezosos arborícolas y primates, lo que
sugiere la presencia de extensos bosques. Otras especies conocidas; Steiromys
detentus, Steiromys duplicatus, Steiromys principalis, Steiromys
pseudonectus y Steiromys
segregatus.
Tracheria troyana.
Krapovickas y Nasif, 2011.
|
 |
|
 |
|
Icnofosil de
las patas delanteras del icnoespecie
Tracheria troyana (*). |
|
Posible
aspecto del Dinomydae
que dejo sus huellas en el Mioceno de La Rioja. Por
MarMag.2026 |
Mamífero Roedor. Se
trata de una icnoespecie, conocida a partir de varias huellas
preservadas. Las huellas de la mano carecen de almohadillas y posee
dígitos asimétricos. El habita del roedor que produjo estas icnitas,
corresponde a zonas fluviales o de terrenos bajos inundados. Las
características anatómicas permiten establecer de que se trata de un
Dinomydae. El Pacarana (Dinomys branickii), es a única
especie viviente entre los roedores Dinomydae, bien diversificado y
distribuido durante el Mioceno – Plioceno, ósea, entre 23 y 3
millones de ańos. Una importante cantidad de muestras de Tracheria
troyana, junto a otros organismos que también dejaron sus
huellas, proceden de la Formación Vinchina (Mioceno), Provincia de
La Rioja, Argentina. Los Pobladores de la ciudad de Vińańa,
intentaron preservar las huellas fósiles cuando estaban a punto de
ser dańado durante las explosiones planificadas durante reparaciones
de la carretera de montańa (R76) que cruza el afloramiento. Así
mismo, posteriormente, un mal entendido con autoridades
locales, autoridades provinciales y policiales, termino con varios
paleontólogos detenidos, y las huellas, depositadas en la comisaria
hasta el día de hoy, por la ausencia de un museo en esta localidad.
Recientemente (2017) se escaneo en 3D del material ubicado en la
estación de policía con el fin de documentar con precisión. Junto a
estas huellas, también se reconocieron a Macrauchenichnus isp
(macrauchenido), caviomorfos o tipoterios, huellas tridáctilas de
reiformes, Gruipeda isp (aves playeras) y Venatoripes
riojanus, un perezoso de tamańo medio.
|
 |
Mar entrerriense.
También
llamado mar paranaense o transgresión entrerriense. Fue un
cuerpo marino del pasado que se formó en la mitad norte del
Cono Sur de América del Sur durante una transgresión marina
del Mioceno, sumado a fases tectónicas andinas, ocurrió una
elevación del nivel marino lo que originó que, avanzando
desde el sudeste (Río de la Plata), la línea costera del
océano Atlántico sudoccidental efectuase un corrimiento
profundo hacia el oeste y norte, internándose en las áreas
continentales, por medio de un mar somero con influencia
deltaica a submareal, representada esta última por barreras
arrecífales carbonáticas, acompańadas por un registro
fosilífero típicamente marino (moluscos de mar, tiburones,
ballenas, etc). Hacia el sur, este mar alcanzó a inundar el
nordeste de la Patagonia argentina, correspondiendo a él los
depósitos de sedimentitas de la formación Puerto Madryn
(este de Río Negro y noreste de Chubut) llegando a latitudes
australes del área de Puerto Madryn y península Valdés. |
Ischyrorhynchus
vanbenedeni. Ameghino 1891
|
 |
|
 |
|
Cráneo de
Ischyrorhynchus vanbenedeni exhibido en el Museo de La
Plata. |
|
Ubicación anatómica del
cráneo del delfín de agua dulce Ischyrorhynchus. |
Mamífero Cetáceo.
Los ínidos o iníidos (Iniidae) son una familia de cetáceos
odontocetos que incluye varias especies de delfines de río o de agua
dulce. Todos los platanistoideos presentan largos y finos
hocicos. Tiene molariformes de corona cónica; paladar ancho y ha
sido descubierto en el Mioceno de Entre Ríos. Su vista es muy
limitada (en algunas especies, inexistentes), por lo que el sentido
de la ecolocalización les resulta indispensable a la hora de
alimentarse y nadar. Tenía una aleta dorsal es poco desarrollada.
Tal vez, una de las características más llamativas es, al igual que
en la familia Monodontidae, la capacidad de mover el cuello, a
diferencia de los delfines oceánicos, que poseen las vértebras
cervicales fusionadas. Poseían una dieta más amplia entre los
odontocetos; se alimentaba principalmente de peces. Seguramente
complementaba su dieta con tortugas de río y cangrejos. Se conocen
pocos materiales y los estudios sobre este grupo son muy
reducidos. Su registro fósil se halla a partir del Mioceno tardío en
Argentina, Brasil y Venezuela.
Notocetus vanbenedeni. Moreno,
1892.
 |
|
 |
| Esqueleto de
Notocetus vanbenedeni
en el Museo Paleontológico 'Egidio Feruglio. |
|
Reconstrucción del aspecto de
Notocetus vanbenedeni del Mioceno de
Puerto Madryn.
(*) |
Mamífero Cetáceo.
Los restos fósiles de este delfín que vivió hace 20 millones
de ańos fueron descubiertos en el área del Parque Industrial
Liviano, en inmediaciones del acceso sur de la ciudad de Puerto
Madryn, en Chubut, Argentina, por científicos del Centro Nacional
Patagónico (CENPAT). Se hallaron vértebras, costillas, algunos
dientes y parte del cráneo de Notocetus vanbenedeni.
Los sedimentos donde estaban los restos fósiles pertenecen a la
denominada Formación Gaiman, y los de esta especie de cetáceos son
abundantes en ella. El ejemplar medía alrededor de tres metros de
largo y presenta un largo rostro y mandíbula. Como antecedente de
ejemplares de esta especie en la zona se destaca el hallazgo de un
esqueleto casi completo y articulado en el valle inferior del río
Chubut, cerca de Gaiman. En el pasado este grupo de delfines estuvo
ampliamente distribuido en Nueva Zelandia, en el Norte América, en
Sudamérica y Europa. Los fósiles pertenecen a una familia de
delfines que ya se extinguió, pero está vinculada con especies de
delfines que todavía viven, aunque poseen otras adaptaciones.
Actualmente la familia está representada sólo por dos especies casi
extintas, Platanista gangetica y Platanista indi,
que viven en India y Paquistán, en los ríos Ganges e Indo. La zona
de Puerto Madryn y Península Valdés, conjuntamente con sus áreas
costeras de influencia del Golfo San José y Golfo Nuevo, son
conocidas y se destacan, principalmente, por la variedad y
exclusividad de su fauna, pero representan también una región de
alto interés desde el punto de vista paleontológico. En los últimos
ańos, su importancia se vio revalorizada por importantes hallazgos
de fósiles tanto de vertebrados como de invertebrados, que
permitieron ampliar el conocimiento de la diversidad biológica en el
área. En este sentido, el personal del Laboratorio de Paleontología
del CENPAT viene realizando un intenso trabajo desde 1989, que
incrementó notablemente el conocimiento de los vertebrados marinos
(mamíferos, peces y aves) del período denominado Terciario del
Atlántico Sudoccidental.
Aondelphis talen.
Viglino, M. et al, 2018.
|
 |
 |
| Posible
aspecto del cráneo de Aondelphis
talen. (*) |
Aspecto de
Aondelphis talen. (*) |
Mamífero Cetáceo.
Es un delfín pequeńo, de aproximadamente dos metros. Fue
bautizado como Aondelphis talen -que en lengua
tehuelche significa delfín del sur pequeńo- por haber sido hallado
en la provincia de Chubut. Si bien los restos que analizamos son
bastante fragmentarios, los huesos del oído permitieron determinar
que se trata de una especie nueva. El animal estudiado, que habitó
los mares patagónicos hace unos 20 millones de ańos, pertenece a la
superfamilia Platanistoidea, un grupo que fue muy diverso en el
pasado, según se infiere por la gran cantidad de especies fósiles
que se encontraron a lo largo del tiempo y en diferentes lugares del
mundo. Sin embargo, en el presente, este grupo se encuentra
representado por una sola especie, que es Platanista gangetica:
el delfín del río Ganges de la India. Las características anatómicas
del oído que presentaban los restos óseos de este delfín permiten
suponer que Aondelphis talen contaba con la habilidad
de la ecolocalización, una característica fundamental que también se
observa en los delfines actuales. El análisis de los sedimentos en
los que se encontraban depositados los restos va a precisar con
mayor exactitud su antigüedad y aportará datos sobre el ambiente y
la fauna. Asociados a los cetáceos encontramos una gran diversidad
de invertebrados, como moluscos, equinodermos, crustáceos, entre
otro. Existe una diversidad de fauna que habitaba el mismo ambiente
que este delfín y que aporta valiosa información.
Saurocetes
argentinus.
Burmeister 1871.
|
 |
 |
|
Fragmento mandibular. Museo
de La Plata. |
Reconstrucción. (*) |
Mamífero Cetáceo.
Fue un cetáceo odontoceto (delfín con dientes) que habitó los
ambientes fluviales y estuarinos de la Patagonia durante el Mioceno,
hace aproximadamente entre
15
y 10 millones de ańos. Sus restos fósiles fueron hallados
en Argentina y representan uno de los mayores delfines de agua dulce
conocidos del registro fósil sudamericano. Su nombre significa
"ballena lagarto de Argentina", en alusión a su gran tamańo
y aspecto alargado. Este mamífero podía superar
los
5 metros de longitud, dimensiones comparables a las de un
gran tiburón blanco y muy superiores a las de los actuales delfines
de río, como el boto amazónico (Inia)
o el delfín del Ganges (Platanista).
Poseía un cuerpo hidrodinámico, un hocico largo provisto de
numerosos dientes cónicos y robustos, adaptados para capturar presas
resbaladizas. Saurocetes fue un
depredador piscívoro, alimentándose principalmente de peces
de gran tamańo, aunque también pudo consumir crustáceos y otros
vertebrados acuáticos. Su poderosa mandíbula y dentición indican que
ocupaba el nivel más alto de la cadena alimentaria en los sistemas
fluviales donde vivía. Durante el Mioceno, gran parte del norte de
la Patagonia presentaba una extensa red de ríos, lagunas y estuarios
conectados con incursiones marinas temporarias. En ese ambiente
convivió con una rica fauna integrada por tortugas, cocodrilos, aves
acuáticas, peces de gran tamańo y numerosos mamíferos. El hallazgo
de Saurocetes argentinus
aporta valiosa información sobre la evolución de los cetáceos
sudamericanos y demuestra que los grandes depredadores acuáticos
también formaban parte de los ecosistemas continentales de la
Patagonia hace millones de ańos.
Aulophyseter
rionegrensis.
Gondar, 1974.
|
 |
 |
|
Diente de Acrophyseter (*). |
Recreación paleoartistica de
Acrophyseter (*). |
Mamífero Cetáceo.
Es un género extinto de cachalotes del
Mioceno, encontrado en las formaciones de las costas este y oeste de
Norteamérica y en la Patagonia en Sudamérica. Cetáceo odontoceto,
es decir, aquellos que poseen dientes. Es un posible sinónimo de
algunos fósiles asignados al género Orycterocetus, que
estaba relacionado con otras ballenas depredadoras como Acrophyseter y
el gran Livyatan. Aulophyseter alcanzaba
una longitud de 6 metros, con un peso corporal estimado en 1100
kilogramos. Hace unos 15 millones de ańos, en la provincia de Río
Negro, el Océano Atlántico había invadido parte de los que hoy es la
estepa patagónica; este mar poco profundo era de un aspecto
tropical, había corales, peces coloridos, ballenas y eran
frecuentados por focas, lobos marinos y delfines. Recientemente se
encontraron dientes de gran tamańo en la provincia de Río Negro en
el yacimiento Base del Gualicho nos indican que estas bestias eran
grandes predadores que se alimentarían de ballenas y otros
vertebrados que podrían atrapar con estos dientes que alcanzaban
hasta 36 centímetros de largo y son los dientes más largos que se
conocen para un vertebrado. Este género fue encontrado primero en
Perú, después en Chile, y esta es la primera vez que se lo encuentra
en Argentina. Su cráneo medía más de 3 metros de longitud y sus
mandíbulas estaban acompańadas por unos 40 dientes de más de 30
centímetros de largo. Actualmente, los dientes del Levyatan
s.p hallados en Río Negro, se encuentran en el Museo
Municipal de Lamarque y el Museo de la Asociación Paleontológica de
Bariloche. Otras especies relacionadas; Aulophyseter
mediatlanticus Cope, 1895 y Aulophyseter morricei Kellogg,
1927.
Idiorophus
patagonicus. Lydekker, 1894.
|
 |
|
.jpg) |
|
Cráneo y
mandíbula de
Idiorophus
patagonicus. |
|
Aspecto de Idiorophus
patagonicus. Por MarMag.2026 |
Mamífero Cetáceo. Es
una
especie extinta de
cachalote primitivo
(Odontoceti, Physeteroidea) que habitó la costa de la Patagonia
argentina durante el Mioceno temprano.
Fue descrito originalmente a finales del siglo XIX por Richard
Lydekker basándose en un cráneo incompleto, y recientemente se ha
realizado una reevaluación anatómica y taxonómica completa de la
especie. Es uno de los fiseteroideos fósiles más antiguos
conocidos y ha sido objeto de estudios recientes para una
redescripción detallada tras más de un siglo desde su hallazgo
inicial.Se
estima que era un cachalote de tamańo mediano, alcanzando
aproximadamente
cinco metros de longitud.
Los estudios paleobiológicos sugieren que su dieta pudo estar basada
en peces y cefalópodos, similar a los cachalotes actuales de menor
tamańo, a diferencia de otros fiseteroideos del Mioceno que eran
depredadores ápice con dietas especializadas en mamíferos marinos.
Los restos fósiles conocidos provienen principalmente de la
Formación Gaiman en Chubut, Argentina.
|
Prosqualodon australis.
Lydekker
1894.
|
 |
 |
 |
|
Cráneo de Prosqualodon australis
en el Museo de La Plata. |
Fósiles de
Prosqualodon
australis
en el Geoparque Bryn Gwyn de
Gaiman, Chubut. |
Posible aspecto de Prosqualodon australis(*). |
Mamífero Cetáceo.
Este extrańo animal vivió del Oligoceno a
principios del Mioceno en Australia, Nueva Zelanda y América
del Sur, y medía 2,3 metros de longitud. Su familia
inmediata puede haber sido los antepasados de todas las
demás ballenas con dientes. Es probable que se pareciera al
pequeńo delfín moderno, con un hocico largo y estrecho,
provisto de dientes puntiagudos, aptos para atrapar peces.
Pero los dientes eran primitivos, ya que todavía le quedaban
dientes triangulares en la parte posterior de las
mandíbulas, como sucedía en las ballenas más primitivas, los
arqueocetos. El cráneo de Prosqualodon se
había vuelto ligero como consecuencia de varias
modificaciones que el animal experimentó en la parte
frontal. En primer lugar, el cuello se había acortado mucho
y la parte posterior de la cabeza se unía al cuerpo y
necesitaba menos apoyo y protección. En segundo lugar, la
compleja estructura mandibular de las ballenas primitivas se
había simplificado considerablemente, gracias a una
alimentación a base de peces. Y, en tercer lugar, como el
olfato ya no era el sentido fundamental que se utilizaba
para localizar las presas (el sonido había ocupado su
lugar), reduciendo el complejo aparato olfatorio. Las fosas
nasales estaban situadas sobre el techo de la cabeza, entre
las cuencas oculares, donde formaban un espiráculo (como el
de las ballenas actuales). El aire viciado que se acumulaba
durante la inmersión se expulsaba de forma explosiva cuando
el animal salía a la superficie.
|
Cetotherium
moreni.
Lyddeker,
1894. |
|
Cetotherium
Cetotherium Cetotherium Cetotherium Cetotherium Cetotherium
Cetotherium Cetotherium
Cetotherium
Cetotherium |
|


 |
|
|
|
Mamífero Cetáceo.
Este Cetáceo pertenecía a una familia de
ballenas primitivas que evolucionó a fines del Oligoceno y
alcanzó su auge durante el Mioceno, hace unos 15 millones de
ańos. Tenía un parecido sorprendente con la ballena gris
actual del Pacífico Norte, aunque tenía menos de la tercera
parte de su longitud. Es probable que sus placas fuesen
bastante cortas, aunque esto resulta difícil de determinar,
ya que la piel de las ballenas, como los cuernos y los
pelos, no se fosilizan. Sin embargo, los cráneos conservan
las marcas de los vasos sanguíneos que aumentaban a las
ballenas durante su vida, y a partir de estos restos se
pueden averiguar muchas cosas. Cetotherium y sus
familiares eran, probablemente, presas de una especie de
enorme tiburón blanco, Carcharodon, que, a
juzgar por el tamańo de los dientes que a menudo se
encuentran fosilizados, alcanzaba un tamańo similar al de
una ballena pequeńa. Tenía 4 metros de longitud. Otro
cetáceo conocido es Aglaocetus morenii. |
Morenocetus parvus. Cabrera, 1926.
 |
|
 |
|
Cráneo de
Morenocetus
parvus de la colección del Museo de La Plata.
Prensa. |
|
Aspecto en vida de
Morenocetus parvus por el
paleoartista Jorge Gonzalez. |
Mamífero Cetáceo.
Se puede distinguir del otro balaenido
mioceno, Peripolocetus, por tener un proceso
cigomático expandido. La longitud del cuerpo de Morenocetus se
estima en unos 5 metros, alimentándose de pequeńos invertebrados
marinos. El primer balaenido reconocido es el Mioceno temprano Morenocetus
parvus en Argentina. Se describió brevemente a partir de dos
cráneos incompletos, una mandíbula y algunas vértebras cervicales
recogidas de la Formación Gaiman del Mioceno inferior de la
Patagonia. Desde entonces no ha sido revisado, por lo que sigue
siendo un cetáceo fósil frecuentemente citado pero enigmático con
gran potencial para arrojar luz sobre la historia temprana de la
corona Mysticeti. Pero estudios recientes, demuestran que Morenocetus es
el balaenido extinto más antiguo hasta ahora, aunque un espécimen
chaeomysticete de depósitos marinos del Oligoceno tardío en Nueva
Zelanda como un posible representante en la línea evolutiva de los
balaenidos. La optimización de la longitud corporal en nuestra
filogenia de Balaenidae, sugiere que la condición primitiva fue una
longitud corporal relativamente pequeńa representada por Morenocetus,
y que el gigantismo se ha adquirido de forma independiente al menos
dos veces (en Balaena mysticetus y Eubalaena spp.)
con la aparición más temprana de este rasgo en el Plioceno tardío
Mioceno-temprano representado por Eubalaena shinshuensis.
Properiptychus argentinus.
Ameghino 1893. |
|
Acrophoca Acrophoca Acrophoca
Acrophoca Acrophoca Acrophoca Acrophoca Acrophoca Acrophoca
Acrophoca Acrophoca |
|

|
|

|
|
Fragmento mandibular
de la Familia Phocidae (focas). |
|
Reconstrucción de
Acrophoca fischeri, similar a
Properiptychus
del Mioceno de Entre Ríos y Puerto Madryn |
Mamífero
Carnívoro. Los fócidos o focas verdaderas
(Phocidae) son una familia de mamíferos pinnípedos marinos adaptados
a vivir en medios acuáticos la mayor parte del tiempo. Las focas
habitan las regiones costeras de buena parte del globo, con la
excepción de las zonas tropicales. Tienen cuerpos alargados y
fusiformes, adaptados a la natación; las extremidades anteriores son
cortas y aplanadas, mejor preparadas para su uso como aletas que
para el desplazamiento en tierra, aunque algunas especies pueden
moverse a gran velocidad empleando movimientos de reptación. Las
aletas posteriores adoptan una posición fija hacia atrás, y no
pueden retraerse. Gervais y Ameghino (1880) dieron a conocer un
supuesto Otariidae, procedente del “Patagónico” de las barrancas del
Paraná (Fm. Paraná o Ituzaingó) identificado como Arctophoca
fischeri. Posteriormente, Florentino Ameghino (1893, 1897)
describió el pinípedo Properiptychus argentinus. El
material tipo de P. argentinus y otros restos fueron
reestudiados por Muizon y Bond (1982), quienes lo reinterpretaron
como un Phocidae Monachinae. Las piezas fósiles, poseen una edad
aproximada de 23 a 7 millones de ańos. Por las características de
los sedimentos hallados, se los reconoce como pertenecientes al
Terciario tardío (Mioceno tardio-Plioceno temprano). Provienen de la
Formación Ituzaingó, mayoritariamente estéril desde el punto de
vista paleontológico, a excepción de su base que ha dado una rica
fauna continental, conocida como "Mesopotamiense" o Conglomerado
fosilífero. Dicha formación se apoya, en discordancia erosiva, sobre
la Formación Paraná, representada por sedimentos marinos del "Mar
Paranense", que cubrió esta región a mediados y fines del Mioceno.
Durante esa época comienza la sedimentación del "Paranense",
rellenando la gran cuenca pampeana. En el área de Península Valdés
afloran sedimentitas marinas del Terciario superior
correspondientes, principalmente, a la Formación Puerto Madryn
(Mioceno medio, 14 millones de ańos) y en menor medida a la
Formación Gaiman (Mioceno inferior, 23 millones de ańos), que aflora
sólo en el Istmo Ameghino. De los niveles inferiores de la misma se
exhumó, en el área del Golfo San José, un esqueleto parcial y
articulado de un pinípedo de la Familia Phocidae (focas). Este
material representa el registro más antiguo y más austral de una
foca fósil para el Hemisferio Sur y una de las más antiguas del
mundo. El ejemplar se presenta algo incompleto pero en muy buen
estado de preservación, faltándole el cráneo, las vértebras del
cuello y parte de los miembros posteriores. Una característica a
destacar de la fina preservación de este material fue la
conservación del contenido estomacal, lo que ha permitido por
primera vez tener la oportunidad de conocer los hábitos alimenticios
de una foca extinguida. Los resultados obtenidos han sido
sorprendentes ya que un primer análisis sistemático indica que esta
foca probablemente pertenezca a la subfamilia Phocinae o sea a las
“focas del norte”. Por lo tanto las implicancias filogenéticas y
biogeográficas son muy amplias.
Metaxyterium s.p
Reinhart, 1976.
|
 |
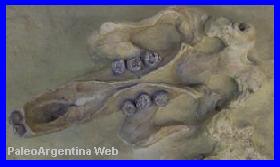 |
 |
|
Esqueletos de Sirenidos.
Ilustrativos. (*). |
Cráneo de
Metaxytherium.
Ilustrativo. Tomado de GAMPS |
Posible aspecto de
Metaxytherium. (*). |
Mamífero Sirenido.
Es un género extinto de mamíferos sirenios de la
familia Dugongidae encontrado fósil en el Caribe, así como en
diversos yacimientos a lo largo de África, Asia, Europa,
Norteamérica y Sudamérica. Los sirenios fósiles de Argentina son
conocidos desde el Mioceno tardío en las formaciones Paraná e
Ituzaingó en la provincia de Entre Ríos. Los sirenios son un orden
de mamíferos placentarios con cinco especies vivientes. Pertenecen
al superorden de los afroterios; entre los animales actuales los
elefantes son sus parientes más cercanos. Son los únicos mamíferos
marinos herbívoros (de ahí que sean llamados "vacas marinas"). Junto
con los cetáceos, los sirenios son los únicos mamíferos adaptados
completamente a la vida acuática. Son superficialmente parecidos a
los pinnípedos (focas y morsas, por ejemplo), con los que no guardan
un parentesco evolutivo; se diferencian de éstos, entre otras cosas,
por el tipo de alimentación (los pinnípedos son carnívoros mientras
que los sirenios son herbívoros) y porque los sirenios son animales
totalmente acuáticos, nunca dejan el agua, ni siquiera para parir.
Son animales grandes y pesados, y tienen movimientos lentos. Viven
en aguas tropicales, en las costas, estuarios e incluso ríos. Además
de los cráneos usualmente más o menos completos de este animal se
han encontrado muchas costillas sólidas para permitir que el animal
permaneciera bajo el agua más tiempo y con menos gasto de energía
durante sus períodos de alimentación a base de algas, que forman
praderas submarinas. Durante el Mioceno Metaxytherium fue
atacado a menudo por el famoso tiburón gigante Carcharodon
megalodon o megalodón. Se ha descubierto el fósil de una
costilla que presenta una serie de perforaciones paralelas
equidistantes entre sí, como un peine: en las costillas quedaron las
marcas de los dientes del megalodón, algunas de las cuales se
originan en la parte superior de los fósiles, mientras que otros
sólo se presentan en la parte convexa, a medida que la influencia de
la mordedura iba en aumento. Otro género conocido de sirénido en el
Plioceno de Argentina es Dioplotherium (Cozzuol, 1996)
y Ribodon limbatus (Ameghino, 1883).
Helianthochelys redondita. Sterli, J. et al,
2025.
|
 |
|
 |
| Vista interna
del caparazón de
Helianthochelys redondita, en el MEF. |
|
Aspecto de la
tortuga marina
Helianthochelys redondita. (*). |
Tortuga
Marina. La Patagonia argentina sigue develando tesoros
paleontológicos de sus entrańas prehistóricas. Esta vez, una nueva
especie de tortuga marina gigante de unos 20 millones de ańos. Según
los expertos, es uno de los registros más completos de este linaje
en el mundo y el más austral conocido hasta ahora para el período
Mioceno. La especie hallada en Chubut, a unos 40 kilómetros de
Trelew, está descrita sobre un único ejemplar, representado por un
caparazón casi completo, con fragmentos de cráneo, vértebras,
pelvis, costillas y la cintura escapula. La especie en cuestión, que
pertenece a una familia llamada dermoquélidas, fue bautizada
Helianthochelys redondita. El nombre proviene de los
términos griegos helios (sol) y anthos (flor), en referencia a la
disposición de los osículos -huesos muy pequeńos- del caparazón, que
recuerdan a la forma de un girasol. Y “redondita” hace alusión a la
localidad de Estancia La Redonda Chica, donde se hallaron los
restos. El ejemplar es un individuo muy viejo y que habría alcanzado
dimensiones similares a las tortugas laúd actuales, con un caparazón
de más de 2 metros de longitud. El registro fósil de este grupo,
cuentan los expertos, es fragmentario y desigual, con apariciones
que se remontan al Cretácico Tardío en Asia y Europa. En Sudamérica,
hasta ahora sólo se habían encontrado restos aislados en Perú y
Argentina. Es por esto que el hallazgo de Helianthochelys
redondita constituye el registro más austral y uno de los
más completos conocidos de este linaje. En estas latitudes, entre
los 23 y los 13 millones de ańos atrás, el clima era más cálido que
el actual. Incluso tenemos registros de más de 40 millones de ańos
que muestran a dermoquélidos llegando hasta la Antártida. Hace unos
20 millones de ańos, hubo una ingresión marina que cubrió la zona
oriental de Chubut. La tortuga Helianthochelys redondita
fue hallada en sedimentos marinos acumulados durante esa ingresión
|
Carcarocles megalodon.
Agassiz, 1843. |
|
Carcarocles megalodon Carcarocles megalodon Carcarocles
megalodon Carcarocles megalodon Carcarocles megalodon |
|

|

|

|
|
Mandíbula de
Megalodon recreada a tamańo natural en el Museo
Paleontológico de Bariloche. |
Diente del
tiburón Carcarocles megalodon, común del Mioceno de
Patagonia y Entre Ríos, Argentina. (*) |
Reconstrucción
del gigantesco Carcarocles megalodon.
(*) |
Pez Selaceo.
Fue encontrado cerca del río Paraná, en Entre Ríos,
donde penetraba el océano Atlántico hace más de seis millones de
ańos. Podía alcanzar un tamańo semejante al tiburón blanco actual y
poseía dientes con serruchos ligeramente pronunciados. El
investigador del Museo de La Plata y Conicet, Alberto Luís Cione,
fue el responsable de este hallazgo cerca de la ciudad Villa General
San Martín. El surgimiento del gran tiburón blanco (Carcharodon
carcharias) ha sido ampliamente debatido entre los
paleontólogos. Sabemos que el origen de estos antecesores inmediatos
del tiburón blanco se da en el Pacífico, porque hay registros en
Perú y Chile, como también en Norteamérica, pero ésta es la primera
vez que se lo detecta en el océano Atlántico. Hay dos hipótesis
sobre cómo este ejemplar hallado pudo haber llegado a la zona donde
hoy está el río Paraná. Una posibilidad es que haya pasado al océano
Atlántico por el área actualmente ocupada por Panamá, porque
Centroamérica y Sudamérica estaban separadas durante el Mioceno
tardío y había una vía de paso entre ambos océanos para los animales
marinos. El itsmo se comenzó a completar hace unos tres millones de
ańos, permitiendo la migración de animales terrestres y el
denominado gran intercambio biótico interamericano. Como hace seis
millones de ańos el nivel del mar era más alto, fue posible que este
tiburón de dientes semi-aserrados nadara hasta donde hoy está el río
Paraná, porque en esa época la llanura pampeana y parte de Entre
Ríos estaban cubiertas en gran medida por aguas saladas. Sin
embargo, también es posible que el ancestro del Carcharodon
carcharias haya cruzado al Atlántico por el estrecho de
Drake, porque esos mares no eran tan fríos en esa época. A través de
ellos, se pudo reconstruir un linaje de tres especies hasta llegar
al tiburón blanco: entre el Isidus plicatilis, que es
la forma más antigua y poseía dientes lisos; una forma intermedia,
que tenía dientes semiaserrados, que se formó en el océano Pacífico,
hace unos siete millones de ańos, y de alguna manera llegó a cruzar
al Atlántico; y, por último, el Carcharodon carcharias, que
es el tiburón blanco tal como lo conocemos ahora. Otro género
conocido; Oxyrhina patagonica.
Carcardum s.p.
Cione, 2012.
|
 |
|
 |
|
Dientes de Carcadum sp
publicados en los medios de comunicación. |
|
Reconstrucción. (*) |
Pez Selaceo.
Fue encontrado cerca del río Paraná, en Entre
Ríos, donde penetraba el océano Atlántico hace
más de seis millones de ańos. Podía alcanzar un
tamańo semejante al tiburón blanco actual y
poseía dientes con serruchos ligeramente
pronunciados.
El investigador del Museo de La Plata y Conicet,
Alberto Luís Cione, responsable de este hallazgo
cerca de la ciudad Villa General San Martín.
El surgimiento del gran tiburón blanco (Carcharodon
carcharias)
ha sido ampliamente debatido entre los
paleontólogos.
Sabemos que el origen de estos
antecesores inmediatos del tiburón blanco se da
en el Pacífico,
porque hay registros en Perú y Chile, como
también en Norteamérica, pero ésta es la primera
vez que se lo detecta en el océano Atlántico.
Hay dos hipótesis sobre cómo este ejemplar
hallado pudo haber llegado a la zona donde hoy
está el río Paraná. Una posibilidad es que haya
pasado al océano Atlántico por el área
actualmente ocupada por Panamá, porque
Centroamérica y Sudamérica estaban
separadas durante el Mioceno tardío y había una
vía de paso entre ambos océanos para los
animales marinos. El itsmo se comenzó a
completar hace unos tres millones de ańos,
permitiendo la migración de animales terrestres
y el denominado gran intercambio biótico
interamericano. Como hace seis millones de ańos
el nivel del mar era más alto, fue posible que
este tiburón de dientes semi-aserrados nadara
hasta donde hoy está el río Paraná, porque en
esa época la llanura pampeana y parte de Entre
Ríos estaban cubiertas en gran medida por aguas
saladas. Sin embargo, también es posible que el
ancestro del
Carcharodon carcharias
haya cruzado al
Atlántico por el estrecho de Drake, porque esos
mares no eran tan fríos en esa época.
A través de ellos, se pudo reconstruir un linaje
de tres especies hasta llegar al tiburón blanco:
entre el
Isidus plicatilis,
que es la forma más antigua y poseía dientes
lisos; una forma intermedia, que tenía dientes
semiaserrados, que se formó en el océano
Pacífico, hace unos siete millones de ańos, y de
alguna manera llegó a cruzar al Atlántico; y,
por último, el
Carcharodon
carcharias,
que es el tiburón blanco tal como lo conocemos
ahora. Otrogenero conocido;
Oxyrhina patagonica.
Hemipristis serra.
Agassiz 1843
|
 |
|
 |
|
Algunos dientes de
Hemipristis serra. (*). |
|
Aspecto de
Hemipristis (*). |
Pez Selaceo.
Es una especie de tiburones de la
familia Hemigaleidae, del orden
Carcharhiniformes, muy parecido al cazón
dientudo actual. Este género fue descrito por
Louis Agassiz en 1843. Hemipristis posee
dos tipos distintos de dientes en cada sección
de su mandíbula. Los que están en la mandíbula
superior actúan como cuchillos, cortando la
carne de la presa, mientras que los puntiagudos
actúan como tenedores, ya que lanzan y sujetan a
la presa. La fauna de elasmobranquios conocida
de la Formación Paraná en la provincia de Entre
Ríos es muy abundante. Se ha encontrado
numerosos dientes atribuibles a Hemipristis
serra, asociados a otros peces miocenos
como Carcharias acutissima, Carcharhinus sp,
Physogaleus aduncus, Heterodontus sp, Squatina
sp, Isurus hastalis y Carcharocles
megalodon. También se lo ha registrado
en formación Puerto Madryn, (Mioceno medio
marino), de Península Valdés en la Patagonia Argentina.
Pogonias sp.
Lacepčde,
1801
|
 |
 |
|
Fragmento de maxilar de
Pogonias sp. Museo Paleontológico de Diamante, Entre
Ríos. |
Aspecto de Pogonias sp.
Por MarMag.2026 |
Pez
esciénido. Se denomina mar
entrerriense, también llamado mar entrerriano, mar paranaense, mar
de Bravard o transgresión entrerriense, a un cuerpo marino del
pasado que se formó en la mitad norte del Cono Sur de América del
Sur durante una transgresión marina del Mioceno. Pogonias era
un pez de agua salobre. La Formación Paraná aflora en las barrancas
paranaenses de Entre Ríos en una angosta lonja paralela al río
Paraná y orientada de norte a sur. Actualmente este género está
representado por la corvina negra (Pogonias cromis),
similar a su pariente, la corvina roja (Sciaenops ocellatus).
Aunque muchos especimenes se encuentran en el rango de 2 a14
kilogramos, la corvina negra es bien conocida por ser la más grande
de la familia de las corvinas, con algunos ejemplares superando los
50 kilogramos y una longitud de 1,70 metro. Su dentadura es
redondeada y tienen fuertes mandíbulas capaces de triturar ostras y
otras conchas. Pertenece a la familia Sciaenidae (esciénidos). Los
Sciaenidae tienen una aleta dorsal larga cerca de la cola.
Genidens ancestralis.
Bogan, S. & Agnolin, F. 2011.
|
 |
|
 |
|
Neurocráneo
de
Genidens ancestralis
en
el Museo de Paleontología de Bariloche.
|
|
Aspecto del
primitivo bagre de mar
Genidens ancestralis.
Por MarMag.2026 |
Pez.
Es la única especie sólo conocida por el
registro fósil de las que componen el género de peces silúridos
marinos Genidens de la familia Ariidae, los que son
popularmente conocidos como bagres de mar. Sus restos fueron
colectados en el norte de la Patagonia Argentina. El holotipo es un
neurocráneo relacionado con un aparato de Weber, las subsecuentes 3
primeras vértebras precaudales, una placa nucal y un fragmento basal
de su espina dorsal, además de elementos de la cintura pectoral y
del esplacnocráneo, todos pertenecen a un mismo individuo. La
localidad donde fue exhumado es el Puesto Picavea, situado en la
región del Gran Bajo del Gualicho, una depresión endorreica ubicada
en el área septentrional de la Patagonia esteparia, en el
centro-norte de la provincia de Río Negro, considerada como
perteneciente al Mioceno Temprano alto a Mioceno Medio bajo. Dicho
Miembro litológicamente destaca por poseer afloramientos de
areniscas, coquinas y pelitas, lo cual se ha referido a un ambiente
transgresivo de aguas marinas.
Phractocephalus yaguaron.
Bogan, S. & Agnolin, F. 2019.
|
 |
|
 |
|
Fragmento de cráneo de Phractocephalus yaguaron. |
|
Aspecto de Phractocephalus yaguaron. Por MarMag.2026 |
Pez.
El río Paraná es el mayor curso de agua de la
Argentina y uno de los más grandes de América del Sur. Su cuenca
tiene una larguísima historia que se remonta a varios millones de
ańos antes del presente. Actualmente se encuentra bien separada de
su vecina del Norte, la super cuenca del río Amazonas. Pero esto no
siempre fue así, y los fósiles de peces son muy importantes para
entender las antiguas conexiones entre las cuencas de los grandes
ríos Sudamericanos. Los investigadores Sergio Bogan y Federico
Agnolín de la Fundación Azara, la Universidad Maimónides y el Museo
Argentino de Ciencias Naturales "Bernardino Rivadavia", dieron a
conocer una nueva especie extinta, pariente del gigante bagre de
cola roja que actualmente vive en los ríos Orinoco y Amazonas. Los
fósiles fueron originalmente hallados en las barrancas del río
Paraná en la provincia de Entre Ríos y tienen 10 millones de ańos de
antigüedad. Los restos del cráneo de este pez son tan grandes y
gruesos que se encontraban hacía ańos en las colecciones del Museo
Argentino de Ciencias Naturales identificados erróneamente como los
restos de un colosal cocodrilo. Los restos conservados indican que
este bagre gigante habría superado los dos metros de longitud y
probablemente más de 70 kilogramos de peso.
Acanthicus.
Agassiz in Spix & Agassiz, 1829.
|
 |
|
 |
|
Fragmento de
cráneo de Acanthicus. Imagen de prensa. |
|
Aspecto del
genero Acanthicus (*): |
Pez.
Los Siuriformes son un grupo de peces
increíblemente diversos que incluye especies conocidas
coloquialmente como bagres, moncholos, surubíes, yuskas, cascarudos
y viejas del agua. Estas últimas forman parte de la familia de los
loricáridos, por su aspecto acorazado y prehistórico. Pese a esta
enorme diversidad, poco es lo que se sabe sobre su historia
evolutiva y los fósiles de viejas del agua son aún poco conocidos.
Es por eso que investigadores se vieron sorprendidos cuando
encontraron grandes restos del esqueleto de gigantescas viejas del
agua que habían pasado desapercibidos. Estos restos proceden de las
barrancas del río Paraná en la provincia de Entre Ríos y tienen una
edad cercana a los 10 millones de ańos de antigüedad. Luego de un
análisis descubrieron que los restos pertenecían a una vieja del
agua gigante llamada Acanthicus (palabra griega que
significa espinoso). Este tipo de pez vive hoy en día en el norte de
Sudamérica, específicamente en la cuenca del Orinoco y en el
Amazonas y es la primera vez que sus restos son descubiertos en el
Paraná. Dentro de toda la diversidad para esta familia de peces, Acanthicus incluye
a la especie viviente de mayor tamańo de viejas del agua, que puede
superar fácilmente el metro de longitud. Se alimentan especialmente
de algas y corteza de ramas sumergidas en el agua. El tamańo de los
restos fósiles encontrados indica que la especie extinta del Paraná
habría sido aún de mayores proporciones. El hallazgo fue realizado
por Sergio Bogan y Federico Agnolin, investigadores de la Fundación
Azara, la Universidad Maimónides, del Museo Argentino de Ciencias
Naturales "Bernardino Rivadavia" y el CONICET. Este nuevo
descubrimiento se suma a otros dados a conocer recientemente como el
de los enormes bagres Brachyplatystoma elbakyani y Phractocephalus
yaguaron, peces gigantes emparentados con los que hoy en día
viven en los grandes ríos del norte sudamericano.
Megapiranha paranensis.
Cione et al. 2009.
Pez.
Hace unos 10 millones de ańos los ríos de la
Mesopotamia Argentina, estaban habitados por una megapirańa, un pez
carnívoro de más de un metro de largo, era algo que se suponía desde
hace tiempo. Sin embargo, hasta el momento no había pruebas de su
existencia. Fue un investigador quien las halló revisando antiguas
cajas con fósiles indiferenciados en los depósitos del Museo de
Ciencias Naturales de La Plata. La reciente publicación de su
hallazgo despertó gran interés, tanto en el ámbito de la
paleontología como en el de la divulgación científica. Revistas como
la National Geographic se hicieron eco del descubrimiento del doctor
Alberto Cione, paleontólogo del Museo de La Plata e investigador del
Conicet. Este material aclara el salto evolutivo que separa a las
pirańas actuales con el pacú, un pariente herbívoro mucho más
primitivo. Aunque se conoce ahora, la historia del descubrimiento se
remonta a fines de 1980, cuando Cione, a cargo de la colección de
peces de la división de Paleontología de Vertebrados, revisaba
antiguas cajas con material fósil sin identificar. Esas piezas
habían sido desenterradas de las barrancas del Río Paraná, cerca de
la localidad de Villa Urquiza, hacia el 1900, y desde entonces
permanecían guardadas. Ancestro directo de las actuales pirańas,
aunque de un tamańo cinco veces mayor, las megapirańas habrían
habitado los ríos, y quizás también las lagunas, de la mesopotamia
hace entre 8 y 15 millones de ańos. Nadie está seguro, sin embargo,
de qué se alimentaban exactamente. Es posible que su dieta fuera
diversa, como la de las pirańas actuales, que, si bien son
eminentemente carnívoras, también comen plantas y frutos. Lo que sí
se sabe es que coexistían con enormes bagres, delfines de agua
dulce, tortugas y cocodrilos de muy diversos tipos, incluso uno
labial de rostro alargado; toda una fauna que se extendía por una
amplia región geográfica hasta el Amazonas. Pese a que las únicas
pruebas de su existencia se hallaron en nuestro territorio, es muy
probable que las megapirańas hayan llegado a habitar también gran
parte de los ríos de Sudamérica. Por entonces las temperaturas
globales eran mucho más altas que ahora, y las cuencas del Paraná y
el Amazonas no estaban, como hoy, aisladas una de otra.
Salminus noriegai.
Cione y Azpelicueta, 2013.
 |
 |
 |
|
Cráneo y Mandíbula. (*) |
Cráneo y Mandíbula. (*) |
Aspecto en vida.
Por MarMag.2026 |
Pez. La nueva especie fue
bautizada como Salminus noriegai, en reconocimiento al
paleontólogo Jorge Ignacio Noriega, que se especializa en el estudio
de aves y fue quien encontró este cráneo de 9 centímetros de
longitud al noreste de la ciudad de Paraná, en la localidad Toma
Vieja. Paleontólogos argentinos rescataron un ejemplar que pudo
haber superado los 50 centímetros, aunque lo más importante no es su
talla, sino que permitió conocer el pasado de estos peces tan
importantes dentro de la fauna de los ríos sudamericanos. El estado
de conservación de la cabeza hallada es muy bueno y, por eso,
permitió hacer un estudio minucioso y poder identificar a este
dorado como una nueva especie. El dorado ancestral se diferencia por
la forma de los huesos en sus mejillas y por la longitud de los
huesos que bordean su boca, como el maxilar y el premaxilar; además,
la órbita ocular es proporcionalmente alargada. Por otra parte, se
observó que sus dientes tienen una forma muy peculiar, símil a un
ocho en la zona que se aferra a la mandíbula. Esta es una cualidad
que comparten con los dorados actuales, pero que nunca se había
descrito hasta ahora, siquiera en los vivientes”, contó el
especialista en peces. Actualmente, viven cuatro especies de dorado
en América del Sur. Es uno de los peces de agua dulce más
importantes de Argentina y Sudamérica; se lo encuentra también en
Brasil y en Colombia, por ejemplo, pero hasta ahora nunca había
aparecido en el registro fósil. En la época inmediatamente anterior
a que viviera este dorado, cuyo cuerpo midió cerca de 60
centímetros, el océano Atlántico ingresaba hasta la provincia de
Entre Ríos, formando un gran golfo. Luego, el mar se retiró y se
formó una serie de ríos. Es por ello que en dicho yacimiento se
encuentran especies de agua dulce y también de agua salada; además,
también se descubren huesos y dientes de los mamíferos que vivían a
orillas de los ríos. De los fósiles encontrados en la formación
fluvial, el Salminus noriegai está entre los de mejor
conservación, por lo que se pudo estudiar tanto los huesos externos
como internos de la cabeza y compararlo con los dorados actuales.
Sanjuanableps calingasta. Bogan, Contreras, Agnolin,
Tomassini y Peralta, 2018.
|
 |
|
 |
|
Fósil de
Sanjuanableps,
recuperado en la zona del Geoparque
Puchuzúm,
quebrada La Candelaria, San Juan. |
|
Posible aspecto de
Sanjuanableps.
Madrecita de agua.
Por MarMag.2026 |
Pez. Los anablépidos,
también conocidos como madrecitas de agua, son una familia de peces
sudamericanos cuya evolución está aún por ser bien
comprendida. Hasta ahora, los especímenes fósiles más antiguos que
se conocen de este grupo provienen únicamente del oeste de la
Argentina y están representados por tres especies muy distintas a
las vivientes. Recientemente, en el departamento sanjuanino de
Calingasta, un equipo conformado por investigadores del CONICET en
el Museo Argentino de Ciencias Naturales “Bernardino Rivadavia” (MACN,
CONICET) y el Instituto Geológico del Sur (INGEOSUR, CONICET- UNS),
junto a colegas de la Universidad de Nacional de San Juan (UNSJ) y
de la Fundación Azara, hallaron en estado fósil varios ejemplares de
lo que resultó ser una nueve especie de anablépido que bautizaron
como Sanjuanableps
calingasta, que vivió entre
hace 9 y 7 millones de ańos durante el Mioceno Tardío. El lugar de
hallazgo constituye una ventana al pasado donde se preservaron hasta
los más finos detalles de decenas de ejemplares de esta especie. El
registro excepcional de estos especímenes permitió el reconocimiento
de individuos masculinos y femeninos, adultos y juveniles. Los
investigadores creen que estos peces habitaron en lagos ubicados en
grandes ambientes áridos de lo que hoy es el territorio argentino.
Probablemente conformaban cardúmenes compuestos solo por individuos
de esta especie, dado que hasta el momento no se registraron otras
especies en los niveles fosilíferos donde fueron descubiertos.
Tenían un claro dimorfismo sexual, donde los machos eran
notablemente más pequeńos que las hembras y contaban con la aleta
anal conformando un órgano copulador muy evidente llamado gonopodio.
Asimismo consideran este aporte en función de la posibilidad de
tener un mayor conocimiento sobre la diversificación de los peces
sudamericanos.
Pindocarpon
chichinalensis.
Muci, Passalia, e Iglesias. 2026.
|
 |
|
 |
|
Frutos fósiles de
Pindocarpon chichinalensis. |
|
Aspecto de
Pindocarpon
chichinalensis, mientras
un perezoso trepa la palmera en la antigua selva patagonica. |
Paleobotanica.
Es una especie extinta de palmera descrita a partir de numerosos
frutos fósiles hallados en sedimentos del Mioceno temprano de la
Formación Chichinales, en el Área Natural Protegida Paso Córdoba,
provincia de Río Negro, Argentina. Vivió hace aproximadamente
20 millones de ańos,
cuando el norte de la Patagonia presentaba un clima subtropical
mucho más cálido y húmedo que el actual. Los fósiles corresponden a
más de treinta frutos excepcionalmente preservados, estudiados
mediante cortes petrográficos, microscopía óptica, microscopía de
epifluorescencia y microtomografía computada de rayos X. Estas
investigaciones permitieron reconocer un nuevo género y especie,
asignados a la subtribu Attaleinae
(tribu Cocoseae), el mismo
grupo al que pertenecen actualmente palmeras sudamericanas como el
Pindó (Syagrus romanzoffiana) y, de manera más
lejana, el Cocotero (Cocos nucifera). Los frutos
presentan las características típicas de las palmeras cocoseas,
entre ellas los tres poros de germinación ubicados en la base del
fruto, además de una anatomía interna excepcionalmente conservada.
La estructura de la pared del fruto muestra rasgos únicos que
sugieren un estrecho parentesco con las actuales palmeras del género
Syagrus, ampliamente distribuidas en Sudamérica. La
gran abundancia de frutos y semillas fósiles recuperados, que
alcanzan cientos e incluso miles de ejemplares distribuidos en
distintos niveles sedimentarios, indica que durante el Mioceno
temprano existieron extensos
palmares en esta región patagónica. Estos bosques
probablemente formaban parte de un paisaje abierto semejante a las
sabanas subtropicales modernas, donde las palmeras constituían
pequeńos bosques intercalados con áreas de vegetación más baja. El
yacimiento de Paso Córdoba conserva una de las paleofloras más
importantes del Mioceno de Sudamérica, junto con abundantes fósiles
de mamíferos, aves, reptiles y plantas. La presencia de
Pindocarpon chichinalensis constituye el primer
registro fósil inequívoco de la subtribu Attaleinae en el sur de
Sudamérica y uno de los frutos fósiles de palmera mejor preservados
conocidos a nivel mundial. Su descubrimiento aporta información
fundamental sobre la evolución de las palmeras, la historia de las
floras sudamericanas y las condiciones climáticas que dominaron el
norte de la Patagonia durante el Cenozoico temprano.
Ostrea patagonica.
d´Orbigny,
1842.
|
 |
|
 |
|
Ostrea maxima,
colectada durante la construcción del Aeropuerto en la Isla
Martín García en 1962 por Carlos Capelli. |
|
Ostrea patagonica,
procedentes del Terciario del Cerro Gualicho
(Oligoceno-Mioceno).
Colección
Museo Municipal Punta Hermengo de Miramar. |
Invertebrado.
Es
un género moluscos bivalvos marinos del orden Ostreoida, conocidos
popularmente como ostras. Poseen dos valvas casi circulares y
desiguales. Durante el Mioceno una gran parte de la Argentina estuvo
cubierta por un mar conocido ampliamente en la literatura geológica
como "mar entrerriense o paranense". Los espesores más potentes
corresponden a los depósitos del subsuelo del ámbito Chaco-paranense,
mientras que secuencias que no superan los 200 metros de espesor,
afloran en Patagonia nororiental (Formación Puerto Madryn) y en el
este de la provincia de Entre Ríos, donde aflora la delgada
secuencia de la Formación Paraná. La característica más notoria de
todos estos depósitos lo constituye la abundante y diversa fauna de
moluscos restringida a las facies marinas de las formaciones Paraná
y Puerto Madryn y a las del subsuelo de la provincia de Buenos
Aires.
Esta intercalación marina, visible en los potentes acantilados, se
caracteriza por grandes concentraciones de valvas de Ostrea
patagonica, que ofrecen una excelente oportunidad para los
estudios tafonómicos e icnológicos.
La gran marca semilunar es donde se inserta el poderoso
músculo abductor. Son bastante comunes de encontrar en sedimentos
terciarios en las costas argentinas, con variable tamańo. Especies
relacionadas; Ostrea alvarezii y
Ostrea maxima.
Monophoraster darwini. Desor, 1847.
 |
 |
 |
|
Vista oral.
Escudo de Mar, un equinodermo marino, común en sedimentos
Miocenos de Patagonia y Entre Ríos. (*) |
Vista aboral
Monophoraster , es un equinodermo común en el Mioceno.
Península Valdés. Col Museo de Miramar. |
Aspecto de un equinodermo
viviente (*). |
Invertebrado.
Es un equinodermo
marino del Mioceno tardío, de Punta Pirámides, Provincia de Chubut, Argentina. La
estrechez de las zonas interambulacral en la superficie oral y en
el ambitus es muy característico y se ve sólo en Eoscutella y Iheringiella. Monophoraster difiere
de esos taxones en tener un lunule anal. Es muy cerca de Amplaster diferentes en
ser mucho menos transversalmente alargado, con una, más periproct de
hendidura más pequeńa que se encuentra en el extremo posterior de
los pétalos posteriores. Una discusión detallada de este género está
dada por Mooi et al. (2000). Las primeras menciones sobre la
existencia estos mares son de índole paleontológica y datan desde
mediados del siglo diecinueve. Fueron efectuadas por d’Orbigny
(1842), quien halló moluscos marinos en los alrededores de la ciudad
de Paraná y en la desembocadura del río Negro. Unos ańos más tarde,
Darwin (1846) encontró la misma fauna en la península Valdés,
quedando así establecida la presencia de estos depósitos en la
Patagonia septentrional y en la provincia de Entre Ríos, los que
constituyeron, por otra parte, las primeras citas del Terciario
marino en la Argentina. Otra especie conocida es; Iheringiella
patagonesis.
Ophiocrossota kollembergorum.
Caviglia, Martínez & del Río, 2007.
|
 |
|
 |
|
Ophiocrossota kollembergorum, hallado en Punta
Maqueda. Formación Chenque, en la provincia de Chubut. (*) |
|
Aspecto de
un Ophiuroidea.
(*) |
Invertebrado.
Los ofiuroideos son una clase de equinodermos).
Presentan simetría pentarradial y tienen un aspecto parecido a las
estrellas de mar (asteroideos), con cinco brazos que salen de un
disco central. Seguramente, ofiuroideos y asteroideos son grupos
hermanos. Un fósil muy interesante corresponde justamente a Ophiocrossota
kollembergorum (Ophiuroidea), Punta Maqueda. Formación
Chenque , en la provincia de Chubut. Tienen el cuerpo pequeńo y
aplanado formado por un disco redondeado por cinco brazos
articulados muy finos y largos que surgen bruscamente del disco
central y pueden estar ramificados. La boca está compuesta por cinco
placas móviles que actúan como mandíbulas. Carecen de ano, por lo
que su tubo digestivo es incompleto. Los materiales no digeridos se
expulsan por la boca. El esqueleto es de gran similitud al de las
estrellas de mar, salvo por la presencia de vértebras esqueléticas
en los brazos, que reducen u obliteran completamente la cavidad
celomática.
Chaceon peruvianus.
d'Orbigny, 1842
Crustáceo. Es un
género de cangrejos marinos. Vivía en aguas
templadas, poco profundas y cerca de la costa. Al igual que en sus
parientes actuales, usaba sus pinzas para separar los trozos de
comida (invertebrados y pequeńos vertebrados) al momento de
alimentarse. Este espécimen fue hallado junto a varios cangrejos y
otros invertebrados en la zona costera de Península Valdés, Patagonia
Argentina. Los sedimentos donde se encontraron los fósiles contienen
ceniza volcánica. Aunque los volcanes estaban a más de 650
kilómetros de la
costa atlántica de donde vivían estos animales, hay estudios que
sugieren que los crustáceos fueron víctimas de afecciones
respiratorias debido al efecto de las cenizas volcánicas en el
tejido blando de su sistema respiratorio.
Éste es un cangrejo fósil cuya taxonomía ha sido
cambiada numerosas veces en diferentes géneros. Su caparazón podía
llegar a medir hasta treinta centímetros de anchura y diecisiete de
longitud y era redondeado, con cinco dientes poco marcados a cada
lado de los ojos, en el borde anterior. Las pinzas eran más cortas
que las demás patas, de dedos suavemente curvados y notablemente
dentados. El segundo par de patas era muy ligeramente más corto y
fino que los tercer, cuarto y quinto pares, los cuales eran más o
menos idénticos en tamańo y proporción. Vivió durante el Eoceno medio, desde hace
cuarenta y nueve millones de ańos, aunque durante mucho tiempo se
había pensado que era propio del Mioceno. Se distribuía por el cono
sur de América, en la Patagonia. Vivía en las aguas profundas, los
juveniles vivían en aguas más profundas que los adultos, y dentro de
éstos, los machos vivían en aguas más profundas que las hembras. Era
carnívoro, alimentándose de moluscos y gusanos del lecho marino. Los
juveniles de esta especie muestran muchas características en común
con los adultos de Proterocarcinus latus. Otros cangrejos
extintos de la Patagonia
son
Archaeogeryon peruvianus y
Archaeogeryon patagonicus.
Otros vertebrados del
Mioceno Argentino:
Mamíferos: Colpodon
burmeisteri, Microbiotherium, Sparassocynus, Proeutatus, Peltocoelus,
Asterostemma, Analcitherium, Neonematherium, Cardiatherium,
Anchimys, Kiyutherium, Proheptoconus, Mixotoxodon, Gyrinodon,
Tetramerorhinus prosistens,
Tetramerorhinus cingulatum,
Picturotherium
migueli, Lambdaconus lacerum,
Heptaconus
obcallatus,
, Asmodeus, Acrotypotherium, Typotheriopsis, Pachyrukhos,
Raulringueletia, Tremacyllus y Pontoplanodes.
Aves: Arthrodytes grandis, Paraptenodytes
antarcticus, Cladornis, Apterodytes, Physornis, Brortornis,
Liornis y Eucallornis, Peces:
Oxyrhina, Lamna, Odontaspis, Notidanus, Scapanorhynchus, Corax,
Hemipristis, Sphirna, Galeocerdoy y Synechodus.
|
(*) Imagen de autor u origen desconocido.
Ver sobre nuestras
Políticas de uso de imágenes.
|
|
Bibliografía Utilizada.
ALBINO, A.M. 1996 b. Snakes from the Miocene of Patagonia
(Argentina). Part I: The Booidea. Neues Jahrbuch für
Geologie und Paläontologie, Abhandlungen 199: 417-434.
AMEGHINO, F. 1888. Rápidas
diagnosis de mamíferos fósiles nuevos de la República
Argentina. Buenos Aires, Obras Completas, 5:471-480.
AMEGHINO, F. 1889.
Contribución al conocimiento de los mamíferos fósiles de la
República Argentina. Actas de la Academia Nacional de
Ciencias de Córdoba, 6:1-1027.
BENTON, M. (1992).
Dinosaurios y otros animales prehistóricos. Ediciones
Lrousse Argentina S.AI.C.
BRAVARD, A. 1858. Monografía
de los terrenos marinos terciarios del Paraná. Imprenta del
Registro Oficial 107 pp. Paraná. (Reimpresión del Congreso
de la Nación 1995)
Bogan, S. & F. L. Agnolin (2011). Descripción
de una nueva especie de bagre marino fósil (Teleostei,
Siluriformes, Ariidae) del Mioceno de la provincia de Río
Negro, Argentina.
Pap. Avulsos Zool. 51(25):373-382.
BORDAS, A.F. 1939. Diagnosis
sobre algunos mamíferos de las capas con Colpodon del valle
del río Chubut (República Argentina). Physis 14 (46):
413-433.
CERDEŃO, E.
2000. Los mamíferos de la Formación Arroyo Chasicó (Mioceno
superior), provincia de Buenos Aires, del Museo "J. C.
Moyano" de Mendoza.
Ameghiniana 37: 503-508.
CIONE, A. L,
AZPELICUETA, M.M., BOND, M., CARLINI, A., CASCIOTTA, J.R.,
COZZUOL, M.A., FUENTE, M. DE LA, GASPARINI, Z., GOIN, F.,
NORIEGA, J., SCILLATO-YANÉ, G.J., SOIBELZON, L., TONNI, E.P.,
VERZI, D., VUCETICH, M.G. 2000.
The Miocene vertebrates from Paraná, eastern
Argentina. In: Aceńolaza, F.G. & R. Herbst (eds.).
El Neógeno de
Argentina. Serie Correlación Geológica 14: 191-237.
CIONE, A.L. Y TONNI, E.P.
1995. Bioestratigrafía y cronología del Cenozoico superior
de la Región Pampeana. En: M.T. Alberdi, G. Leone y E.P.
Tonni (eds.), Evolución biológica y climática de la Región
Pampeana durante los últimos cinco millones de ańos, pp.
49-74.
CIONE, A.L.; TONNI, E.P.;
BARGO, S.; BOND, M.; CANDELA, A.M.; CARLINI, A.A.; DESCHAMPS,
C.M.; DOZO, M.T.; ESTEBAN, G.; GOIN, F.J.; MONTALVO, C.I.;
NASIF, N.; NORIEGA, J.I.; ORTIZ JAUREGUIZAR, E.; PASCUAL,
R.; PRADO, J.L.; REGUERO, M.A.; SCILLATO-YANÉ, G.J.;
SOIBELZON, L.; VERZI, D.H.; VIEYTES, E.C.; VIZCAÍNO, S.F. &
VUCETICH, M.G. 2007. Mamíferos continentales del Mioceno
tardío a la actualidad en la Argentina: cincuenta ańos de
estudios. Asociación Paleontológica Argentina, Publicación
Especial, Ameghiniana 50ş aniversario, 11:257-278.
CHIAPPE, L. 1991. Fossil birds from the Miocene Pinturas
Formation of Southern Argentina. Journal of Vertebrate
Palaeontology 11, supplement (3): 21A-22A.
COZZUOL, M.A.
1993. Mamíferos acuáticos del Mioceno medio y tardío de
Argentina. Sistemática, evolución y biogeografía. Tesis
doctoral, Universidad Nacional de La Plata, Facultad de
Ciencias Naturales y Museo de La Plata, inédita.
DE IULLIS, G.,
RÉ, G.H. y VIZCAÍNO, S.F. 2004.
The Toro Negro megatheriinae (Mammalia,
Xenarthra): a new species of Pyramiodontherium and a review
of Plesiomegatherium. Journal of Vertebrate Paleontology 24:
214-227.
Dozo, M.T. &
Vera, B. (2010).
«First skull and associated postcranial bones of
Macraucheniidae (Mammalia, Litopterna) from the Deseadan
SALMA (late Oligocene) of Cabeza Blanca (Chubut,
Argentina)». Journal of Vertebrate Paleontology 30
(6): 1818-1826.
ESTEBAN, G.I. Y
NASIF, N.L. 1999. Mamíferos fósiles de la Formación
Chiquimil (Mioceno tardío), provincia de Catamarca,
Argentina. Bioestratigrafía. 15° Jornadas Argentinas de
Paleontología de Vertebrados, Ameghiniana Suplemento
Resúmenes 36: 11R.
ESTEBAN, G.I.,
NASIF, N.L.y MONTALVO, C.I. 2001. Nuevos registros de
Dasypodidae (Xenarthra) del Mioceno tardío de la provincia
de La Pampa, Argentina. Revista Espańola de Paleontología
16: 77-87.
FORASIEPI, A. M., GOIN, F. J.
AND MARTINO, V. 2003. Una nueva especie de Lycopsis (Metatheria,
Prothylacyninae) de la Formación Arroyo Chasicó (Mioceno
Tardío) de la provincia de Buenos Aires.
Ameghiniana 40(2):249-253
GASPARINI, Z. 1973. Revisión
de los Crocodilia (Reptilia) fósiles del territorio
Argentino. Su evolución, sus relaciones filogenéticas, su
clasificación y sus implicancias estratigráficas. Tesis
Doctoral 314 Facultad de Ciencias Naturales La Plata: 1-169.
GEORGIEFF, S.M., HERBST, R., ESTEBAN, G.I. y
NASIF., N. 2004.
Análisis
paleoambiental y registro paleontológico de la Formación
Desencuentro (Mioceno Superior), Alto de San Nicolás, La
Rioja, Argentina. Ameghiniana 41: 45-56.
GOIN, F.J., MONTALVO,C.I. y VISCONTI, G.
2000.
Los
marsupiales (Mammalia) del Mioceno Superior de la Formación
Cerro Azul (provincia de La Pampa, Argentina). Estudios
Geológicos 56: 101-126.
GUILLERMO
W. ROUGIER, JOHN R. WIBLE, ROBIN M. D. BECK AND SEBASTIAN
APESTEGUÍA (2012). «The Miocene mammal Necrolestes
demonstrates the survival of a Mesozoic nontherian lineage
into the late Cenozoic of South America». Proceedings of the
National Academy of Sciences of the United States of
America. in press.
KRAMARZ, A.G. 2001A. Estudio
de la fauna de roedores de la Formación Pinturas, Mioceno
Medio temprano de la Provincia de Santa Cruz. Tesis Doctoral
(Inédito), Universidad de Buenos Aires, Facultad de Ciencias
Exactas y Naturales: 183 p.
KRAGLIEVICH, L (1931):
Contribución al conocimiento de las aves fósiles de la época
araucoentrerriana. Physis 10: 304-315.
LEONE Y E.P. TONNI (eds.),
Evolución biológica y climática de la Región Pampeana
durante los últimos cinco millones de ańos: 259-275. Madrid.
MAGNUSSEN
SAFFER, M. (2013). Aves gigantes representativas y extintas
de América. Paleo, Revista Argentina de Paleontología.
Boletín Paleontológico. Ańo 11. 85: 17-23.
MAGNUSSEN
SAFFER, M. (2013). Características de los reptiles mas
conocidos en el Mioceno de Argentina. Paleo, Revista
Argentina de Paleontología. Boletín Paleontológico. Ańo 11.
86:28-33.
MAGNUSSEN
SAFFER, M. (2013). Mamíferos marinos característicos del
Mioceno de la Republica Argentina. Paleo, Revista Argentina
de Paleontología. Boletín Paleontológico. Ańo 11. 87: 17-22.
MOOI,
R., MARTINEZ, S. & PARMA, S. G. 2000.
Phylogenetic systematics of tertiary monophorasterid sand
dollars (Clypeasteroida: Echinoida) from South America.
Journal of Paleontology 74, 263-281.
NOVAS, F. 2006. Buenos Aires
hace un millón de ańos. Editorial Siglo XXI, Ciencia que
Ladra. Serie Mayor.
PARDIŃAS, U.F.J. 1991.
Primer registro de primates y otros vertebrados para la
Formación Collón Cura (Mioceno medio) del Neuquén,
Argentina. Ameghiniana 28 (1-2): 197-199.
PASCUAL, R., ORTEGA
HINOJOSA, E., GONDAR, D.E. y TONNI, E. 1966. Paleontografía
Bonaerense, Fascículo IV, Vertebrata (Ed. Borrello, A.V.).
Comisión de Investigaciones Científicas de la Provincia de
Buenos Aires, La Plata, 202 pp., CI láminas.
PASCUAL, R.; BONDESIO, P.;
VUCETICH, M.G.; SCILLATO YANÉ, G.J.; BOND, M.; TONNI, E.P.
1984. Vertebrados fósiles cenozoicos. In Congreso Geológico
Argentino, No. 9, Relatorio 2: 436-461. Buenos Aires.
PATTERSON, B.
& KRAGLIEVICH, L (1960): Sistemática y nomenclatura de las
aves fororracoideas del Plioceno Argentino. Publicación del
Museo Municipal Ciencias Naturales
y Tradicionales de Mar del Plata 1: 1-51
ROVERETO, C. 1914. Los
estratos Araucanos y sus fósiles. Anales del Museo
Nacional de Historia Natural de Buenos Aires 25: 1-247.
SCILLATO YANÉ,
G.J., CARLINI, A.A. y VIZCAÍNO, S.F. 1987. Nuevo
Nothrotheriinae (Edentata, Tardigrada) de edad chasiquense
(Mioceno Tardío) del sur de la provincia de Buenos Aires
(Argentina). Ameghiniana 24: 211-215.
Soibelzon, Leopoldo Héctor; Bond, Mariano; Revisión de los
carnívoros (Mammalia, carnivora) acuáticos y continentales
del Mioceno de la Mesopotamia argentina; Asociación
Paleontológica Argentina; Publicación Electrónica de la
Asociación Paleontológica Argentina; 14; 1; 11-2013; 164-172
SORIA, M.F., JR. 2001. Los
Proterotheriidae (Litopterna, Mammalia), sistemática, origen
y filogenia. Monografías del Museo Argentino de Ciencias
Naturales (1): 1-167.
TAMBUSSI, C.;
AGNOLIN, F.; COZZUOL, M. 2003. Un nuevo depredador en el
elenco de aves de los ecosistemas miocénicos patagónicos. In
Jornadas Argentinas
de Paleontología Vertebrados, No. 19, Resúmenes: p. 30.
Buenos Aires.
Tambussi, C. P.; De Mendoza, R.; Degrange, F. J.; Picasso,
M. B. (2012). «Flexibility along the Neck of the Neogene
Terror Bird Andalgalornis steulleti (Aves
Phorusrhacidae)». En Evans, Alistair Robert. PLoS ONE
7 (5): e37701.
TAUBER, A.; GARRIDO, A.;
TAMBUSSI, C.; KRAMARZ, A.; SALDIVIA, S. 1999. Una nueva
localidad con vertebrados continentales del Mioceno,
Departamento de Picún Leufú, Neuquén. Ameghiniana 36,
suplemento (4): p. 22R.
TAUBER, A.A. 1997.
Paleoecología de la Formación Santa Cruz (Mioceno Inferior)
en el extremo sudeste de la Patagonia. Ameghiniana
34:517-529.
Verónica Krapovickas, Nahuel A. Muńoz, Pablo
Alonso Muruaga, Patricia
L. Ciccioli, Sergio Marenssi & Sergio F. Vizcaíno (2017):
Tracheria Troyana (Krapovickas and Nasif, 2011):
Redefinition, Environmental Distribution, and Heritage
Conservation, Ichnos.
VUCETICH, M.G.; MAZZONI,
M.M.; PARDIŃAS, U.F.J. 1993. Los roedores de la Formación
Collón Cura (Mioceno medio), y la ignimbrita Pilcaniyeu.
Cańadón del Tordillo. Neuquén. Ameghiniana 30 (4): 361-381.
VUCETICH,
M.G. y VERZI, D. 1995. Los Roedores Caviomorfos. En: M.T.
Alberdi, G. Leone y E.P. Tonni (eds.), Evolución biológica y
climática de la Región Pampeana durante los últimos cinco
millones de ańos, pp. 213-225.
Madrid.
WICKER, R. 1984. Beobachtungen bei
mehrjähriger Zucht von Phrynops geoffroanus geoffroanus
(Schweigger, 1812) (Testudines: Chelidae).
Salamandra
20(4): 185-191.
Ver mas
bibliografía utilizada para hacer la presente Pagina;
AQUÍ.
Ver bibliografía de divulgación sugerida;
AQUÍ. |
|
|
|
  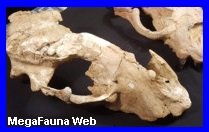 |
|
 Ver
otros continentes en el mismo Periodo Geológico Ver
otros continentes en el mismo Periodo Geológico |
|














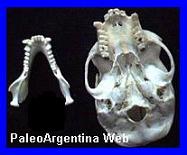
.jpg)
.jpg)
.jpg)






.jpg)
.jpg)