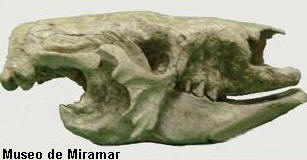
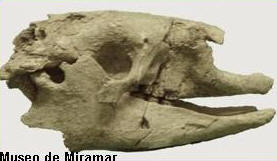
Un perezoso gigante junto a su
cría. Algunos datos sobre su estudió.
Mariano Magnussen Saffer. Museo Municipal Punta
Hermengo de Miramar. Fundación Argentavis. Grupo
Paleo.
marianomagnussen@yahoo.com.ar
Tomado de;
Magnussen Saffer,
Mariano (2014).
Un perezoso gigante junto a su cría. Algunos datos sobre su
estudió. Paleo Revista
Argentina de Paleontología. Boletín Paleontológico. Ańo XII. 107: 19-22.
Ilustraciones
y fotografías de Daniel Boh, Paleoartista,
museólogo y titular del Museo Municipal Punta
Hermengo de Miramar.
Recientemente, y por medio de un trabajo
científico, se dieron a conocer estudios
realizados a un ejemplar adulto y un juvenil de
Scelidotherium leptopcephalum en el
interior de una paleocueva colapsada del
Pleistoceno superior, en la cual, se realizaron
observaciones tafonomicas sobre ambos individuos
y el contexto en donde y como fue hallado.
El material procede de la localidad balnearia
de Mar del Sud, Partido de General Alvarado,
Provincia de Buenos Aires, Argentina, y
recuperado, restaurado, estudiado y depositado
en el Museo Municipal “Punta Hermengo” de la
ciudad de Miramar.
La Tafonomía es la parte de la Paleontología que estudia los procesos de
fosilización y la formación de los yacimientos
de fósiles. Se puede ayudar de disciplinas como
la Ecología, la Geoquímica, la Sedimentología,
etc. Junto a la Paleobiología y la Biocronología,
la Tafonomía es una de las tres divisiones de la
Paleontología.
|
 |
Los fósiles que podemos encontrar en un
estrato no tienen por qué provenir de
organismos que vivieron juntos, que
compartieron el mismo ambiente e
interactuaron, ni siquiera ser de la
misma edad geológica.
Los estudios tafonómicos nos pueden
permitir analizar la historia y
procedencia de los diferentes elementos
de una asociación fósil, para ello hay
que tener en cuenta todos los parámetros
observables posibles, características de
la matriz rocosa, rellenos,
alteraciones, encostramientos,
abrasiones, erosiones, mineralizaciones,
deformación y rotura, posición y
orientación, disgregación de partes,
selección de tamańos, colonización, etc.
Cuando en un mismo estrato se pueden
distinguir agrupaciones de fósiles que
comparten algunos de estos factores en
común se pueden establecer «familias»
tafonómicas con los restos que han
compartido una historia común, lo que
indicaría una evolución tafonómica
compleja, y hay que estudiar, por tanto,
cada agrupación separadamente. |
Como en otras disciplinas, lo complejo del análisis
tafonómico, exige un lenguaje específico, preciso,
que exprese inequívocamete los conceptos
involucrados y necesarios para las interpretaciones
paleobiológicas, paleoecológicas, bioestratigráficas
y paleogeográficas. Por otra parte, los fósiles no
son los organismos que vivieron en el pasado, aunque
fueran producidos directa o indirectamente por
ellos, y destacar esta obviedad es crucial para
evitar errores y falsas interpretaciones del
registro fósil.
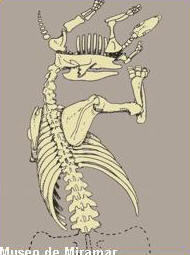
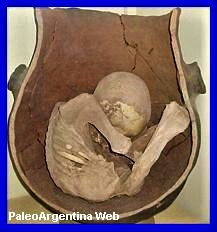
En
las tareas de campo realizadas por el museo
miramarense, se recuperaron del espécimen adulto
prácticamente todo el esqueleto, pero carece de los
miembros traseros debido a la erosión, en cambio del
juvenil se encuentra completo. Este ultimo, se
encontraba apoyado sobre la cabeza del adulto.
El análisis del contexto sedimentario y los
atributos tafonómicos observados permitieron tener
una amplia información del material exhumado. Los
datos aquí reunidos son congruentes con los
antecedentes disponibles, indicando la ocupación de
grandes madrigueras
ha
sido principalmente postulada como producto de la
actividad cavadora de los grandes Mylodontidae
(perezosos gigantes como Lestodon,
Glosotherium y Scelidotherium).
 |
El paquete sedimentario de la paleocueva se
encuentraba compuesto por limos arcillosos
pardo-rojizos y limos arenosos de igual
coloración, generalmente macizo y
litológicamente similares al sedimento
hospedante.
Sus límites pueden ser distinguidos a pesar
de la poca diferencia estructural,
granulométrica y cromática entre el relleno
y el sedimento hospedante, características
previamente citadas para la identificación
de este tipo de icnofósiles. |
Se
considera como sedimento de relleno aquel, que
ingreso a la cueva, tanto por el derrumbe, o por
medio de agua u otro agente. Así mismo, se considera
sedimento hospedante aquel que se encuentra por
debajo, por encima o laterales de la cueva, por lo
general mas consistente.
No
se observan indicios de meteorización (expuesto al
sol, lluvia o agentes ambiéntales) o fragmentación
ósea, surcos o marcas de actividad depredadora,
desmembramiento postmortem o transporte (desplazados
por agua, otros animales etc).
Los
restos postcraneanos (esqueleto) se encuentran
articulados y asociados a los elementos craneanos.
Ambos animales se hallaban un una posible cámara,
sin evidencia de galerías debido a la erosión.
El diámetro de la estructura que contenía el
material, de 1.80 a 2,20 mtrs
correspondiendo al morfotipo ya conocidos de
Scelidotheriinae. La asociación de estos
individuos permite postular que pertenecen al mismo
grupo familiar (madre y cría), y que encontraron la
muerte al derrumbarse la estructura sobre ellos,
seńala el estudio.
Los
primeros restos fósiles atribuidos a Scelidotherium
fueron descubiertos por Charles Darwin en su viaje
del Beagle en la localidad de Bahía Blanca. Basados
en restos del cráneo, vértebras y costillas, Richard
Owen lo bautizo como Scelidotherium en 1840.
|
|
De tamańo menor y del grupo de los
Xenarthros (perezosos) mencionados
anteriormente fueron los mamíferos mas
comunes del paisaje del Pleistoceno, ya que
sus restos fósiles son muy abundantes en los
depósitos sedimentarios de este periodo.
Tenía 1,7 metros de alto y 3,5 de largo. Su
peso estimado es de una tonelada. El cráneo
es alargado, bajo y estrecho, con un rostro
proyectado hacia adelante.
Su alimentación consistía de vegetales que
hallaba en áreas secas. Investigadores del
Museo de La Plata, habían realizado estudios
biomecánicos en los huesos de las patas
delanteras de Scelidotherium,
concluyendo que estos extintos animales
estaban plenamente capacitados para emplear
sus fornidos brazos para cavar. La robustez
de cada uno de sus huesos, así como las
amplias manos en forma de palas, son
coherente con este comportamiento de
superexcavadores. |
Recientemente se pudo saber por varios hallazgos
realizados por el Museo de Ciencias Naturales de Mar
del Plata que estos enormes mamíferos cavaban
madrigueras, las cuales realizaban para refugiarse,
o bien para invernar, ya que es posible que tuvieran
un metabolismo muy bajo, logrando una temperatura
constante de 15° c en su interior.
Una
de ellas media 40 metros de largo con un diámetro de
1,8 de donde se rescataron numerosos moldes de las
garras entre otras evidencias. El autor encontró en
Agosto de 2002 en el Pleistoceno de Miramar una
Paleocueva similar y de menor espectaculidad, la
cual se encuentra en estudio.
El
género Scelidotherium, básicamente
diferenciable de Scelidodon por la presencia
de ciertos caracteres más derivados a nivel de los
miembros y cráneo, cuenta con al menos dos especies
pleistocenas: S. floweri y S.
leptocephalum. Las especies de este género
habitaron en espacios abiertos, parcialmente
arbolados, de climas templados.
Durante el Pleistoceno más tardío se lo encuentra
también en ambientes de altura, compatibles con
condiciones ambientales templado-frías, entre 2.000
y más de 3.000 m, específicamente en las localidades
de Las Juntas, Catamarca y Lauricocha, Perú, aunque
es importante mencionar que en este último sitio
existen evidencias que los restos fueron llevados
allí por el hombre. Geográficamente, se habría
distribuido en los territorios de Argentina, Perú,
Uruguay y Paraguay.
Bibliografía sugerida.
Imbellone, P. & Teruggi, M. 1988. Sedimentación
crotovínica en secuencias cuaternarias bonaerenses.
II Reunión Argentina de Sedimentología, Actas, p.
125-129.
Magnussen Saffer, Mariano (2008). Una cueva
realizada por mamíferos gigantes en el Pleistoceno
de Miramar. Paleo, Boletín Paleontológico. Ańo 6.
Numero 29: 30-34
M. Magnussen Saffer, D. Boh y C. Estarli.
(2014). Asociación de un ejemplar juvenil y uno
adulto de Scelidotherium leptocephalum Owen, 1839 (Xenarthra,
Mylodontidae, Scelidotheriinae ) en una paleocueva
en el Pleistoceno del Partido de General Alvarado.
Aspectos Sistemáticos y Tafonomicos.
XXVIII Jornadas
Argentinas de Paleontología de Vertebrados. Zapala –
Villa El Chocon, Neuquén. Mayo del 2014.
Libro de Resumenes. , p
38.
Scillato-Yané, G.J.; Carlini, A.A.; Vizcaíno, S.F. &
Ortiz Jaureguizar, E.
1995. Los xenartros. In: Alberdi, M.T.; Leone, G. &
Tonni,
E.P. (eds.) Evolución biológica y climática de la
región
pampeana durante los últimos cinco millones de ańos,
Un
ensayo de correlación con el Mediterráneo
occidental.
Museo Nacional de Ciencias Naturales, Madrid,
Monografías, 12:181–209.
Zárate, M.A.; Bargo, M.S.; Vizcaíno, S.F.; Dondas,
A. & Scaglia, O. 1998. Estructuras biogénicas en el
Cenozoico tardío de Mar del Plata (Argentina)
atribuibles a grandes mamíferos. Revista de la
Asociación Argentina de Sedimentología, 5:95-103.
Fósiles marinos en la cima del cerro
Otto.
Por Ari Iglesias. Paleontólogo del INBIOMA (CONICET-UNCOMA).
Algunas fotos han sido procesadas o de archivo para
ser adaptadas al sitio.
Fuente; anbariloche.com.ar
Diversos estudios dan cuenta de que Bariloche estuvo
sumergida en agua salada de mar. La presencia de
caracoles, almejas, erizos y corales respaldan esta
teoría. Un recorrido por la historia.
Es bastante frecuente que barilochenses y turistas
que recorren las montańas y las playas de la región
se acerquen al Museo Paleontológico de Bariloche
consultando por extrańas formas y colores que
hallaron dentro de las rocas.
Muchas de estas rocas revelan la presencia de
caracoles, almejas, erizos y corales que evidencian
que en algún momento, la región de Bariloche estuvo
sumergida en agua salada de mar. Analizando
detenidamente estas formas fósiles dentro de la
roca, los afortunados descubridores asienten con la
evidencia indiscutible de que se trata de organismos
marinos, pero seguidamente se quedan atónitos al
pensar que, para que estos fósiles existieran, el
nivel de mar debería haber pasado por arriba de la
cima del cerro Otto (o lo que es aún más
impresionante que la cima del cerro Otto sea el
fondo de un gran mar).
 |
żEs que el nivel del mar puede haber subido
tanto en períodos de tiempo de hace millones
de ańos, simulando un evento de diluvio
universal? o es que alternativamente, żlos
depósitos marinos del fondo del mar fueron
transportados al lugar que hoy se encuentran
en la cima de una montańa?
Los estudios geológicos y paleontológicos en
la región evidencian que ninguna de estas
dos preguntas es correcta. Varios estudios
sobre el nivel de los mares indican que aún
en aquellos momentos de máximo calor en la
Tierra (cuando no existían glaciares ni
hielo ni siquiera en los polos), todo el
agua en estado líquido en los mares del
mundo, no llegaría a superar en 100 metros
el nivel del mar actual. |
Si consideramos que Bariloche se encuentra a 800
metros sobre el nivel del mar, podemos estar seguros
de que por más catástrofes que puedan ocurrir en el
mundo o que podamos ver en alguna película
taquillera, jamás el mar llegará a Bariloche (al
menos en los próximos millones de ańos hacia
adelante)
Pero entonces, żcómo es que podemos encontrar
fósiles marinos en Bariloche?
Los geólogos y paleontólogos sabemos que los
procesos de cambio de los paisajes pueden tomar
mucho tiempo (millones de ańos). Hoy existen
diversas formas para saber la edad de las rocas,
saber la especie animal que el fósil representa es
una. También se puede saber la edad de formación de
una roca al analizar los elementos radioactivos que
poseen algunos cristales dentro de ellas. Mediante
estos métodos sabemos que las rocas marinas que se
hallan en Bariloche son de un rango de tiempo que
van de los 20 a 30 millones de ańos.
En ocasiones, el movimiento de las placas tectónicas
hace que regiones de grandes proporciones se muevan
a lugares remotos, por ejemplo la India se movió
desde Antártida hasta chocar con Asia, o nosotros
mismos (América del Sur) nos separamos de África a
una velocidad promedio de 3 cm. por ańo (aunque en
un terremoto grande podemos movernos varios metros
en poco tiempo). Pero sabemos que las rocas marinas
en Bariloche siempre estuvieron aquí (no fueron
transportadas grandes distancias).
El estudio de los fósiles marinos en nuestra región,
ha permitido saber que las aguas marinas que bańaban
las costas de Bariloche, en realidad, eran del
océano Pacífico. Las especies de los océanos de uno
y otro lado de Argentina son muy diferentes.
 |
En base a los estudios geológicos también
sabemos que la Cordillera de los Andes para
ese momento del tiempo geológico
(correspondiente a lo que denominamos
Oligoceno Tardío y Mioceno Temprano), no
tenía las elevaciones que hoy conocemos. Y
entre volcanes de pequeńa altura (uno de
ellos el Cerro Leones, en la salida hacia el
Río Limay), el mar ingresaba hasta cubrir la
región de Villa La Angostura, Bariloche, El
Bolsón y llegó a Esquel. |
El estudio paleontológico de fósiles de cangrejos
tipo Paromola y corales solitarios hallados en la
región de el Río Foyel y Península Llao Llao, ha
permitido reconocer especies muy raras de fondos
marinos de alta profundidad. La presencia de estos
organismos permite saber que la región de Bariloche
se encontraba sumergida a más de 200 metros abajo
del nivel del mar (condiciones de oscuridad y
temperaturas frías).
Pero, seguimos sin entender żcómo es que ahora
encontramos las rocas y los fósiles en la cima del
cerro Otto?
Lo que ocurrió posteriormente al gran desarrollo de
un mar epicontinental (por encima del Continente
Sudamericano), fue aún más espectacular. Luego de
depositarse una importante cantidad de sedimentos en
el fondo de una gran cubeta marina (preservando los
fósiles que hoy conocemos), el aumento en la
velocidad de convergencia (choque) entre la placa
tectónica del Pacífico con la Placa Sudamericana,
realizó un fenómeno orogénico de tamańo descomunal.
Se elevan gigantescas montańas desde Tierra del
Fuego hasta Colombia. La región de Bariloche también
lo hace, lo cual provoca un retiro del mar y genera
que lo que antes era el fondo marino, se eleve a más
de 2.000 metros por sobre el nivel de mar y comience
un lento proceso de meteorización y erosión que
genera las montańas. Si bien en términos geológicos
esto pasó rápidamente, en realidad tomó 20 millones
de ańos en dejar las rocas donde hoy las vemos. Las
rocas marinas de la cumbre del Cerro Otto son
evidencia de que ese fondo marino fue sobreelevado
hasta los 1800 metros de altura y posteriormente la
erosión alrededor del cerro produjo la forma que
tiene hoy.
 |
Fenómenos climáticos y de erosión en los
últimos 3 millones de ańos y 10.000 ańos,
generar una acumulación de nieve de
formidables proporciones, en lo que se
conoce como la Edad de Hielo.
Gigantescos glaciares bajaban desde lo más
alto de las montańas produciendo anchos
valles glaciarios en los cuales hoy se
encuentran todos los grandes lagos de
nuestra región. Estos grandes glaciares
arrastraron como monumentales topadoras
todas las rocas que hoy faltan alrededor del
cerro Otto (que antes eran continuas con el
Cerro Ventana y el Catedral). |
Existen evidencias que demuestran que aún por encima
del cerro Catedral ocurrían más de 1000 metros de
estas rocas marinas, pero que los glaciares se las
han llevado en su movimiento continuo hacia la
región del Río Limay y el Río Pichileufu, pero eso
pertenece a otro momento de la historia geológica de
la región de Bariloche.
Algunas curiosidades de la fauna
triásica (Therapsida, Archosauria y Labyrinthodontia)
de Ischigualasto.
Por
Mariano Magnussen Saffer. Presidente de la
Asociación Amigos del Museo Municipal Punta Hermengo.
Grupo Paleo.
marianomagnussen@yahoo.com.ar.
La fauna triásica de Ischigualasto, constituida
principalmente por cinodontes gonfodontes (Familia
Traversodontidae) y rincosaurios, y en la que
abundan también dicinodontes, arcosaurios basales
rauisúquidos y aetosauridos, y algún laberintodonte,
cuenta además con dinosaurios. En ella se ha
comprobado por primera vez y de manera inobjetable,
la asociación de dinosaurios y cinodontes. Sin lugar
a dudas, Ischigualasto encierra valiosa información
sobre la evolución de variados grupos de Therapsida,
Archosauria y Labyrinthodontia, así como también
sobre los antiguos dinosaurios saurisquios conocidos
como Eoraptor lunensis y
Herrerasaurus ischigualastensis y el
ornitisquio más antiguo Pisanosaurus mertii,
que serán mencionados en artículos posteriores.
Ischigualasto, conocido turísticamente como "Valle
de La Luna", fue declarado Parque Provincial por
iniciativa del Gobierno Provincial y de la
Universidad Nacional de San Juan. A principios de
esta década se llevaron a cabo expediciones
conjuntas entre investigadores argentinos y
extranjeros, participando así el Museo de Ciencias
Naturales de la Universidad Nacional de San Juan, el
Museo Argentino de Ciencias Naturales "Bernardino
Rivadavia" y la Universidad de Chicago (Estados
Unidos).
 |
La Formación Ischigualasto es el sector más
bajo de la Hoyada que lleva ese mismo
nombre. Está rodeada por la Formación Los
Rastros al oeste y la base de la Formación
Los Colorados al este. La misma se extiende
hasta la Hoyada del Cerro de Las Lajas,
cercana a la ruta nacional número 40. Cubre
un área trazada por unos 25 km. de largo y 5
km. de ancho. Los afloramientos fosilíferos
están ubicados en el departamento de Valle
Fértil, aproximadamente 70 kilómetros al
noroeste de la cabecera de ese departamento. |
En
escala cronológica esta Formación es más moderna que la
Formación Los Rastos (parte superior del Triásico Medio)
y más antigua que la Formación Los Colorados, ubicándose
en la parte baja del Triásico Superior, de unos 210
millones de ańos de antigüedad.
Tiene entre 500 y 600 metros de espesor y está
conformada por tobas, tufitas, areniscas finas y
limoarcillitas, obteniendo un paisaje donde predominan
por excelencia los tonos grisáceos. Estos estratos
presentan una inclinación aproximada a los 8 grados
respecto de la horizontal, lo cual permite tener acceso
a diferentes niveles.
Durante el Triásico Superior, la actual zona abarcada
por esta Formación correspondió a un paleoambiente
pantanoso, con lluvias de cenizas y material
piroclástico, alternado con depósitos fluviales
arenosos. La coloración grisácea de los sedimentos
contrasta con las capas rojas de la Formación Los
Colorados. El paisaje de Ischigualasto está compuesto
por pequeńas elevaciones, valles, planicies de erosión y
barrancas con distinta pendiente.
La
majestuosa vista panorámica de Ischigualasto, con
caprichosos y fantásticos relictos de erosión, nada
tiene que ver con el paisaje que presentó en aquellas
épocas remotas, cuando tuvo lugar el depósito de los
sedimentos. Se trata de una marcada depresión "hoyada",
en continua erosión, surcada por una cantidad de arroyos
secos que desaguan en épocas de lluvia.
Los
vertebrados triásicos de la Formación Ischigualasto
representativos son: Labyrinthodontia:
Promastodonsaurus belemanni
Bonaparte, 1963 y Pelorocephalus ischigualastensis
Bonaparte, 1975. Therapsida: Ischigualastia
jenseni Cox, 1964. Cynodontia: Exaeretodon
frenguellii Cabrera, 1943. Ischignathus
sudamericanus Bonaparte, 1963.
Chiniquodon
sp. Bonaparte, 1966. Probainognathus sp.
Bonaparte y Crompton 1995. Rynchosauria: Scaphonix
sanjuanensis Sill, 1970. Archosauria:
Saurosuchus galilei Reig, 1959.
Aetosauroides scagliai
Casamiquela, 1960. Venaticosuchus rusconii
Bonaparte, 1970. Trialestes romeri
Reig, 1963 y Proterochampsa barrionuevoi
Reig, 1959.
Gualosuchus reigi.
Las características de este genero son muy parecidas a
aquellas citadas para Chańaresuchus bonapartei.
Su cráneo posee un hocico bajo y alargado, compuesto de
una buena cantidad de dientes cónicos y aserrados para
capturar presas. Es muy probable que pasaría un tiempo
muy importante dentro de los espejos de agua,
alimentándose de peces como el descrito mas abajo, o de
las tortugas acuáticas. El largo de esta especie esta
calculado entre los 1,5 a 2 metros de largo y unos 45
centímetros de altura.
Gracilisuchus stipanicicorum.
De esta especie se conocen varios ejemplares
relativamente completos, caracterizados por presentar un
tarso cocodriloide (de cocodrilo). Su tamańo era de
pequeńas dimensiones, ya que media 70 centímetros de
largo por 30 centímetros de largo. El cráneo de este
pequeńo reptil Arcosaurio es proporcionalmente bajo y
alargado. Su vista dorsal es triangular, con el extremo
anterior muy angosto y la región posterior ancha.
 |
Las grandes orbitas tienen una amplia expresión
dorsal, por lo que los ojos se proyectarían
laterodorsal y anteriormente, tal vez con
considerable superposición de campos visuales.
Su mandíbula es algo mas corta que la longitud
del cráneo, llegando solo a la mitad del pre-maxilar.
Poseía dos vértebras sacras, mientras que su
larga cola poseía unas 45 vértebras de tamańo
decreciente. La extremidad posterior del tarso
es derivado en relación a tecodontes mas
antiguos como Euparkeria,
registrado en el triasico inferior de África.
|
En
Gracilisuchus podemos observar a simple vista
que el miembro anterior es 40% menor que las
extremidades posteriores, lo cual es algo muy frecuente
entre los los Ornitosuquidos. Se trato de un pequeńo
depredador con ciertas habilidades, cuyo alimento
estaría constituido principalmente de insectos y
vertebrados pequeńos. Para culminar, podemos decir que
Gracilisuchus es un interesante ejemplo de
la diversidad de Tecodontes que surgieron o se
diferenciaron durante el triasico medio, como producto
de una gran radiación adaptativa operada en este grupo
de Arcosaurios, y que culminaria con la diferenciación
de ambos ordenes de Dinosaurios (Saurisquia y
Ornitisquios), de los cocodrilos y eventualmente de los
Pterosaurios, que prosperaron a fines del triasico (?) y
principios del Jurasico.
Aetosauroides scagliai.
Los diversos ejemplares disponibles de Aetosauroides
corresponden a especimenes desde unos 80 centímetros
hasta 2,50 metros de largo, en algunos casos con muy
buena información sobre el cráneo y poscraneo, aunque no
totalmente completo. El cráneo de esta especie parece
ser mas primitivo que en los Aetosaurios de Europa y
América del Norte, especialmente por ser mas alargados,
bajo y ramas mandibulares extensas y débiles.
Sus
dientes también son algo mas primitivos, ya que son
pequeńos y agudos en lugar de gruesos y subcilindricos
como en el caso de Typothorax del Triasico
superior de U.S.A. Poseían a lo largo de su columna
vertebral una tipo de coraza, formadas de placas
paramedianas rectangulares, dispuestas transversalmente
en el sector dorsal, y una hilera de placas
cuadrangulares en el borde o costado lateral.
Las
mismas presentan en sus proporciones según correspondan
a la región cervical, dorsal o caudal, pero poseen una
decoración uniforme, constante, de pequeńas fosetas.
Algunos huesos de sus extremidades presentan
características propias de los cocodrilos, demostrando
que se podían mover a cierta velocidad. Esta especie de
la fauna de Ischigualasto, compilo un rol muy importante
en su nicho ecológico, ya sea necrófago u omnívoro -
herbívoro, que no poseía ningún otro Tetrápodo de esa
fauna.
El
Doctor Casamiquela, quien estudio los primeros restos de
Aetosauroides, decidió dedicarle a la especie como "scagliai",
en honor a Galileo Juan Scaglia, ex-director del Museo
de Ciencias Naturales de Mar del Plata, por formar parte
de unos de los primeros grupo de investigadores que
exploraron el "Valle de la Luna" en la Provincia de San
Juan.
Ischigualastia Jensini. Posee un cráneo
voluminoso, con la mandíbula gruesa, masiva, que sugiere
una fuerte capacidad para cortar y triturar vegetales.
Sus ramas mandibulares se encontraban recubierta por
unos fuertes músculos. La parte mas anterior del hocico
era proporcionalmente angosta, lo que sugiere que
seleccionaría de alguna manera los vegetales de que se
alimentaba.
 |
El esqueleto recuperado y montado en el Museo
Pedagógico de la Fundación "Instituto Miguel
Lillo" en la ciudad de Tucumán, nos muestra que
su estructura esqueletaría era muy fuerte,
gruesa, conformando un animal de robustez
comparable o posiblemente mayor a la de un
rinoceronte africano actual. Se destacan las
cortas y gruesas extremidades anteriores, con el
humero en una posición casi horizontal y
dirigida hacia afuera, lo que posibilito a este
animal apoyar su cuerpo sobre cinco dedos muy
fuertes. |
Las
extremidades posteriores eran totalmente lo contrario,
tomando una posición totalmente vertical, lo que hace
pensar a los Paleontólogos que habría sido mas esbelta
en la región posterior y mas voluminosa y pesada en su
mitad anterior, con el cuello corto, grueso y una pesada
cabeza en posición baja, tal vez con algún parecido
general a los búfalos actuales. Procede de los niveles
inferiores de la formación Ischigualasto, ubicado en el
"Valle de la Luna", Provincia de San Juan, Republica
Argentina.
Exaeretodon frenguelli.
Se conocen numerosos restos procedentes de la formación
Ischigualasto, y pertenecen por lo menos a tres o cuatro
especies distintas con características morfológicas
ampliamente diferenciales en las mandíbulas. Los cráneos
coleccionados de este genero oscilan entre 18 y 60
centímetros de largo total.
Observando el cráneo podemos decir que es bastante mas
largo que ancho, con un hocico largo. Posee tres
incisivos en cada pre-maxilar, un canino relativamente
grande y una hilera de pos-caninos. Entre los restos mas
importantes rescatados, podemos mencionar un esqueleto
completo de la sección media de la formación
Ischigualasto, el cual tiene unos 2 metros de largo
(incluido el graneo y la cola que son muy cortas).
Poseía
un tórax voluminoso y huesos de la cintura escapular y
pélvica muy modestos. Estos datos sugieren que era un
animal muy robusto, de patas cortas y gruesas, y de unos
60 centímetros de alto. Era un animal de movimientos
lentos, aunque si era acechado por algún carnívoro podía
ser algo mas ágil pero por breve tiempo. Es una de las
especies mas representativas y abundantes del Triasico
Argentino.
Scaphonyx sanjuanensis.
Se trata de una especie cuyo largo total oscilaba en los
2 metros hasta los 2,5 metros de largo, y una aproximada
de 45 a 50 centímetros. La posición de las extremidades
era primitiva, o sea con el humero y el fémur próximos a
la posición horizontal, lo que contribuía a la poca
altura de los ejemplares.
 |
En la formación Ischigualasto, especialmente en
el terciario inferior, la frecuencia de
Scaphonyx es notable e incluye
ejemplares de diversos tamańos que presentan,
muy probablemente, estadios de desarrollo de
individuos de la misma especie. En el Museo de
Ciencias Naturales de la Universidad Nacional de
San Juan se encuentra montado un magnifico
ejemplar de esta especie. Es indudable que
Scaphonyx sanjuanensis era una
especie totalmente herbívora, y que
eventualmente seria una de las presas de los
grandes carnívoros como Saurosuchus
o aun Herrerasaurus. |
Los
vegetales que probablemente se alimentara este
Rincosaurio es mas difícil estimar. Tal vez la
adaptación de su pico estaba dirigida a arrancar raíces,
o eventualmente para triturar frutos de cicadales según
los autores.
Ischigualasto es un reservorio excepcional de flora y
fauna triásica de ambiente continental, siendo el más
importante en su tipo a nivel mundial, ya que los cuatro
millones de ańos iniciales del Triásico Superior que
están muy bien documentados en los citados
afloramientos, no están registrados, hasta el momento,
en ninguna otra parte del mundo y si lo están han
brindado restos muy incompletos.
Bibliografía Sugerida.
ALCOBER, O.A., 1993. Geología de La Formación
Ischigualasto. Tesis inédita. FCEFN-UNSJ. San Juan.
BONAPARTE, J.F., 1969. Los tetrápodos triásicos de
Argentina. I Simposio de Gondwana, La Estratigrafía del
Gondwana. UNESCO.
BONAPARTE, J.F., 1969. Datos sobre la evolución
paleoecológica en las Formaciones triásicas de
Ischigualasto-Villa Unión. Acta Geológica Lilloana, 10
(9). Tucumán.
BONAPARTE, J.F., 1978. El Mesozoico de América del Sur y
sus tetrápodos. Opera Lilloana, 26: 1-596. Tucumán.
BONAPARTE, J.F., 1996. Dinosaurios de América del Sur.
Museo Argentino de Ciencias Naturales "Bernardino
Rivadavia". Buenos Aires.
CABRERA, A., 1943. El primer hallazgo de terápsidos en
la Argentina. Notas del Museo de La Plata,
Paleontología, 55: 317-331. La Plata, Provincia de
Buenos Aires.
CASAMIQUELA, R., 1960. Noticia preliminar sobre dos
nuevos estagonolepoideos argentinos. Ameghiniana, 2 (1):
3-9. Buenos Aires.
MONETTA, A.M., 1995. Ischigualasto-Talampaya, tiempo de
dinosaurios, time of dinosaurs. Manrique Zago, 200
páginas.
NOVAS, F.E., 1986. Un probable terópodo (Saurischia) de
la Formación Ischigualasto, Triásico Superior, San Juan,
Argentina. IV Congreso Argentino de Paleontología y
Bioestratigrafía, 2: 1-6.
REIG,
O.A., 1961. Acerca de la posición sistemática de la
Familia Rauisuchidae y del género Saurosuchus,
(Reptilia-Thecodontia). Publicación del Museo de
Ciencias Naturales de Mar del Plata, 1 (3): 73-114. Mar
del Plata, Provincia de Buenos Aires.
Los Dinosaurios del
Grupo Neuquén.
Por
Juan
Manuel Morales, Colaborador Centro Paleontológico
Barreales. Neuquén, Argentina. Publicado el Sábado 12 de Abril de 2003.
Contacto
por e-mail:
jamorales@neunet.com.ar
Introducción:
El Grupo Neuquén se le llama a los
estratos rojos que contienen fósiles de dinosaurios y
se encuentran en esta provincia, los cuales ya son
conocidos desde hace muchos ańos en norpatagónia.
Este grupo es uno de los más
importantes del mundo en lo que respecta a fósiles de
dinosaurios y de otros animales del Mesozoico ya sea por
su variedad de especies
como por su importancia paleobiogeográfica.
El Grupo Neuquén se divide en tres
formaciones: Río Limay (350 metros de espesor), Río
Neuquén (150 metros de espesor) y Río Colorado (200
metros de espesor). Estos, a su vez se dividen en
miembros: Mb.Candeleros, Mb. huincul, Mb. Lisandro
(R. Limay), Mb.Portezuelo, Mb. Plottier (R. Neuquén) y Mb. Bajo de la Carpa y Mb.
Anacleto pertenecen a la formación Río Colorado.
Las edades que abarca van desde el
Albiano superior hasta el Campaniano inferior.
Río
Limay.
Andesaurus delgadoi:
Miembro Candeleros, 100 millones de ańos. Fue el primer
dinosaurio encontrado en el Chocón. Este se considera
un titanosaurio primitivo debido a que sus vértebras
caudales anteriores tienen la cara anterior cóncava y
la posterior convexa mientras que las vértebras
caudales medias y posteriores tienen sus ambas caras
planas .También las espinas neurales laminares en las vértebras
caudales y una estructura primitiva llamada hipósfenohipantro
en las vértebras dorsales demuestran que no es un
titanosáurido común del Cretácico superior. Medía 16
metros de longitud y pesaba alrededor de 20 toneladas
Rebbachisaurus tessonei:
Es uno de los saurópodos más completos de Sudamérica.
También es el diplodócido más primitivo encontrado
hasta ahora. Esto esta demostrado debido a caracteres
primitivos como: sus vértebras
cervicales y dorsales no tienen la espina neural
en forma de "v" sino
en forma simple y recta como una "I''. Otra
característica primitiva es la forma curva de sus
dientes a diferencia de los dientes más rectos del
diplodocus.Otra característica muy importante de este
saurópodo es su parentesco con el Rebbachisaurus
garasbae de Marruecos ya que permite suponer una posible
conexión terrestre entre estos dos continentes hace 100
millones de ańo. Rebbachisaurus compartió su entorno
con el Andesaurus y Giganotosaurus. Su medida se calcula
alrededor de los 17 metros y pesaba unas 10 toneladas.
Giganotosaurus carolinii:
Considerado como el mayor depredador del periodo cretácico
con sus casi 14 metros de longitud, con la cabeza a una
altura de 8 metros y con un peso de 10 toneladas
Giganotosaurus es el depredador terrestre más grande
encontrado hasta hoy. Este debía alimentarse de los
saurópodos Rebbachisaurus y Andesaurus con los cuales
convivía diariamente. Giganotosaurus pertenece a la
familia de los carcharodóntidos a la cual pertenece el
gigante carcharodontosaurus saharicus del norte de África
y según estudios muy recientes el Megaraptor
namunhuaiquii (de la formación R. Neuquén y Miembro
Portezuelo). Se cree que Giganotosaurus era un cazador
porque a diferencia de el famoso Tyranosaurus rex los
dientes de este carcharodóntido eran estrechos y
estaban diseńados para arrancar y/o cortar carne, no
como los
del T. rex que eran anchos diseńados para triturar
huesos.
Argentinosaurio huinculensis:
Miembro Huincul, 95 millones de ańos. Titanosaurio de
unos 40 metros de longitud y unos 15 mt. de altura, hoy
considerado como el dinosaurio más grande del mundo. Se
han encontrado muy pocas partes de este titanosaurio,
solo un 10% ha sido rescatado. Pero para tener una idea
del tamańo total de el animal solo hay que imaginar el
tamańo de una vértebra de 1,60 metros de alto y 1,40
metros de ancho. Argentinosaurus vivió en lo que
actualmente es Plaza huincul y Cutral Có.
Anabisetia saldiviai:
Pequeńo dinosaurio terópodo de no más de 2 metros de
longitud y 60 centímetros de altura. El rasgo más
importante de Anabisetia es que permite corroborar la
teoría planteada por Salgado y Coria acerca del origen
de los ornitópodos como Gasparinisaura en territorio
Sudamericano y no como animales procedentes de América
del norte que inmigraron
hacia el sur al formarse el istmo de Panamá.
Esta teoría fue propuesta
junto con el hallazgo del Gasparinisaura (70
millones de ańos), pero la edad de este dinosaurio
coincidía justamente con la formación del istmo de
Panamá (el cual genero un intercambio faunístico de
Sudamérica y Norteamérica) por lo cual se creía que
estos animales habían llegado a
la Patagónia desde América del Norte. Para
comprobar la teoría de Salgado y Coria se necesitaba
hallar un ornitópodo emparentado con Gasparinisaura,
pero de formaciones geológicas más antiguas y esto es
precisamente el Anabisetia saldiviai.
Probablemente este pequeńo dino vivía
en manadas alimentándose de la vegetación que crecía
al nivel del suelo, al igual que Gasparinisaura tenía
un complejo aparato de masticación con unas hileras de apretados
dientes ideales para triturar todo tipo de materia
vegetal. Su única defensa contra los depredadores eran
sus ágiles extremidades posteriores que le permitían a
estos escapar a toda velocidad de sus depredadores.
Río
Neuquén.
Futalongkosaurus dukei:
Miembro Portezuelo. Saurópodo, titanosaurio de 35 o 40
metros de longitud y aproximadamente 15 metros de alto. Se encontró
en el Centro Paleontológico los Barreales en la
provincia de Neuquén. Se ha rescatado 3 ejemplares muy
completos (uno grande y dos pequeńos). A pesar de tener
un tamańo similar al Argentinosaurus no están
estrechamente emparentados entre si. Probablemente
Futalongkosaurus como todos los saurópodos
tenían algún
comportamiento del tipo
gregario que les permitía tener una mayor
seguridad ante los depredadores que compartían su
ambiente como Megarraptor. Estos titanosaurios vivieron
hace unos 87 millones de ańos en lo que era un ambiente
en el cual abundaban lagunas y lagos pequeńos
que contenían gran variedad de fauna y flora.
Megaraptor namunhuaquii:
Dinosaurio terópodo de 6,50 metros aproximadamente. Hasta
hace poco sólo se conocía por una garra de unos 27
centímetros, pero el hallazgo reciente de una mano de
70cm articulada en el Centro Paleontológico los
Barreales nos permiten aprender nuevas características
de este animal: su garra gigante no estaba ubicada en el
pie (similar a los raptores)
como se suponía sino en la mano, esto no solo
cambia de lugar la garra del Megaraptor sino al la de
todos los raptóridos (del pie a la mano) y la otra
novedad es que según indica la anatomía de la mano del
Megaraptor este dinosaurio se incorporaría a la familia
de los carcharodóntidos.
Unenlagia comahuensis:
dinosaurio con características avianas de 2.30 metros
de longitud t 1.20 mt. de alto. La peculiaridad de este
dinosaurio es la forma en que articula el húmero y la
escápula permitiendo que el brazo o proto ala (nombre
que se le da a los brazos del Unenlagia por ser
parecidos a las alas actuales de las aves, pero más
primitivas y que no
servían para volar) se pueda elevar con mayor
amplitud que el de los otros dinosaurios terópodos .
Este movimiento sería muy similar al que realizan las
aves voladoras de la actualidad con sus alas cuando están
volando. Según el descubridor del Unenlagia , el doctor
Fernando Novas, mientras corría este dino daba saltos
cada vez más altos ayudados por los ''aleteos'' que
realizaban sus proto alas. También lo ayudaban a
incrementar su velocidad y a mantener el equilibrio.
Patagonykus puertai:
dinosaurios encontrado a las afueras de Cutral Có con
una edad aproximada de unos 90 millones de ańos. Es un
terópodo aviano de 1.80 metros de longitud muy
emparentados con Alvarezsaurus calvoi (de Neuquén) y
Mononykus olecranus (de Mongolia), juntos forman la
familia Alvarezsauridae. Estos se caracterizan por tener
una garra muy desarrollada en la mano. La anatomía de
el brazo es muy similar a la de los actuales mamíferos
cavadores, pero en casi imposible que los
Alvarezsauridae utilizaran su garra para esto porque las
extremidades anteriores son muy pequeńas en relación a
cuerpo y les resultaría imposible cavar una cueva
relativamente grande. Hasta hoy la función de sus
garras es desconocida, pero quizás sirvió para extraer
raíces o cortezas de las plantas. A pesar de sus
dientes probablemente no comían carne
Río
Colorado.
Patagopteryx deferrariisi:
Ave no voladora encontrada en las bardas de la ciudad de
Neuquén. Esta ave cuenta con unos miembros anteriores
muy reducidos, pero sus patas
son muy robustas y están adaptadas a la carrera,
también fueron utilizadas para la construcción del
nido. Era un ave pequeńa ya que medía aproximadamente
unos 40 cm de longitud y pesaba 3 Kilogramos. Vivió
hace unos 85 millones de ańos.
Alvarezsaurus calvoi:
Dinosaurio terópodo encontrado por el paleontólogo
Jorge Calvo y descripto por
el Doc. José Bonaparte. Este dinosaurio
encontrado en las bardas de Neuquén capital es un
miembro de la familia Alvarezsauridae por lo cual es muy
similar al Patagonykus y al Mononykus.
Sólo se ha rescatado un 40% del
esqueleto total, pero es suficiente como para
describirlo en
su totalidad. La familia Albarezsauridae está muy
emparentada con Archaeopteryx (la ave más primitiva
conocida hallada en
Alemania). Alvarezsaurus vivió hace 85 millones
de ańos, no superaba el metro de longitud y de altura
estaba entre los 70 centímetros.
Velolocisaurus unicus:
dinosaurio terópodo de porte pequeńo que habito
la ciudad de Neuqúen hace unos 85 millones de ańos.
Este dinosaurio esta representado
solo por una extremidad posterior, esta es muy
particular y está muy bien adaptada a la carrera,
cuenta con una anatomía no registrada en otros terópodos
ya que tiene solo un metatarso para sus tres dedos.
Probablemente Velocisaurus era un animal muy agil y de
movimientos ligeros. Se supone que no habría tenido
dietes y que comía insectos y pequeńos animales.
Antarctosaurus wichmannianus:
titanosáurido de 30 metros de longitud aproximadamente.
Vivió en Río Negro y en Neuquén hace un poco más que
80 millones de ańos. Contaba con cráneo corto pero
aplanado en la parte anterior, tenía dientes como
tachuelas que sólo se ubicaban en la parte anterior de
la mandíbula y que utilizaba para arrancar las hojas y
el follaje que deseaba comer. La trituración estaba
destinada a las gastrolitas (o piedras estomacales) como
en la mayoría de los saurópodos, ya que estos
necesitaban comer mucha cantidad de alimento pero sus
mandíbulas no estaban diseńadas para triturar.
Neuquenornirs
volans: ave voladora de la ciudad de Neuquén. Según las
sinapomorfías del húmero, radio, unla, fémur,
tarsometatarso y las vértebras pertenecientes a la región
del tórax, Neuquenornis pertenece a un grupo de aves
extintas llamadas Enantornites. También podemos decir
que según características de el tarsometatarso
Neuquenornis pertenece a la familia del cretácico tardío
llamadas Avisauridae. Se caracteriza por tener rasgos
primitivos en los miembros posteriores y rasgos modernos
en los miembros anteriores y cintura escapular notándose
así la temprana diferenciación de las aves, en el
miembro posterior rasgos primitivos mientras que en el
miembro anterior y cintura escapular sufrieron rápidas
modificaciones adaptadas para el vuelo.
Neuquensaurus australis:
Titanosaurido que rondaba los 9 o 10 metros de longitud.
Fue muy común en patagónia hace unos 85 millones de ańos,
este dino al igual que Saltasaurus estuvo cubierto por
un escudo óseo que probablemente utilizaba como defensa
ante los depredadores de esa época.
Aucasaurus garridoi:
terópodo de 7 metros de longitud, 2 metros de alto y un
peso de 3 toneladas era un cazador activo, veloz y
eficaz. Aucasaurus pertenece a la familia de los
Abelisauridos siendo este uno de los más completos
registrados hasta la fecha. Se hallo en la zona de Auca
Mahuevo (famosa por anteriores hallazgos de huevos fósiles,
de embriones en perfecto estado y hasta piel
embrionaria).Como es frecuente en los Abelisáuridos,
Aucasaurus también poseía algún adorno en su cráneo,
una pequeńa protuberancia ósea sobre los ojos que no
llegaban a ser cuernos.
Titanosaurus araucanikus:
Titanosauridos muy comunes. Estos, al igual que muchos
saurópodos poseían una articulación fuertemente
convexa en las vértebras caudales que le permitían una
mayor movilidad en la cola.
Titanosaurus australis:
Titanosáurido de 8 metros de longitud. Debido a la
cantidad de ejemplares hallados pertenecientes a este género,
se ha podido estudiar con mucha exactitud, también se
han encontrado huevos pertenecientes a esta especie tan
abundante.
|
 |
 |
|
Reconstrucción de Patagopteryx deferrariisi, en la Exhibición del
Museo de Geología y Paleontología del Comahue.
Imagen: Proyecto Dino. |
Reconstrucción de la cabeza de Unenlagia comahuensis. Ilustración de la Pagina
Web Proyecto Dino. |
LECTURAS
SUGERIDAS:
Salgado L.& Pasquali R., ’’El cómo, cuando y
donde de los dinosaurios de la Argentina’’ revista
Ciencia Hoy Volumen
11 - Nş 65- Octubre/Noviembre 2001.
Chiappe
L. M. & Calvo J. O., ’’Neuquenornis volans, a
new late cretaceous bird (enantiornithes: avisauridae)
from Patagonia, Argentina’’. Journal of Vertebrae
Paleontology. June 1994. 14(2): 230-246.
Chiappe
L. M & Calvo J. O.,’’ El primer
enantiornithes (aves) del Cretácico de la Patagonia’’.
VI jorn, Arg, de Paleontología de Vertebrados. San
Juan. Argentina. 1989. pp 19-21.
Bonaparte
J, 1996, ‘’Dinosaurios de América del Sur’’,
Buenos Aires, Museo Argentino de Ciencias Naturales
‘Bernardino Rivadavia’.
Calvo
J. O. & Gazzera C. E. ,‘’Paleoecología
en el sector Inferior del Miembro Candeleros (Fm. Río
Limay, Grupo Neuquén, Cretácico) en el área del lago
Exequiel Ramos Mexía, Prov. del Neuquén, Patagonia,
Argentina’’. VI jorn, Arg, de Paleontología de
Vertebrados. San Juan. Argentina. 1989. pp 3-5
żA
qué época se remonta la presencia humana
en la Argentina?
Por
Gustavo G. Politis - Facultad de Cs. Sociales (Olavarria,
Prov. Buenos Aires, Argentina) Universidad Nacional del
Centro. Publicado originalmente en la revista Ciencia
Hoy Vol1 N° 2 . Ilustrado por PaleoArgentina Web.
Desde una posición extrema que llegó
a sostener el mismo origen de la humanidad en la pampa,
se pasó luego, como contrapartida, a creer que el
poblamiento de América era un fenómeno relativamente
reciente. Hoy, a la luz de nuevos hallazgos arqueológicos,
se ha replanteado también la última postura; en la
Argentina, las evidencias demuestran que en algunos
casos, como el de las llanuras bonaerenses, el hombre
coexistió hace milenios con una fauna extinguida, que
en parte explotó para su supervivencia.
Cuando a fines
del siglo pasado el célebre investigador Florentino Ameghino propuso que la cuna de la humanidad se
encontraba en la pampa argentina, la atención de la
comunidad arqueológica mundial se volcó sobre la región.
żEra posible que, contrariando a la mayoría de los
hallazgos de la época, los fragmentarios cráneos y
otros restos óseos exhumados en la llanura pampeana
pertenecieran a los primeros hombres que poblaron el
planeta? El cúmulo de información expuesta por
Ameghino era importante y mereció una discusión
internacional. De esta manera, en 1910 llegó a la
Argentina el paleoantropólogo (especialista en la
evolución biológica del hombre) Ales Hrdlicka, de la
Smithsonian Institution de Washington; luego de un rápido
análisis de los materiales y de las evidencias
presentadas por el científico local, destruyó las
propuestas del mismo, concluyendo además que no había
razón para creer que el hombre en América del Sur
tuviese más que unos pocos milenios de antigüedad.
Tampoco era probable, según su opinión, que hubiera
convivido con los grandes mamíferos del Pleistoceno.
|
 |
La
contundencia del modelo del prestigioso Hrdlicka,
excelentemente presentado en su EARLY MAN IN SOUTH
AMERICA (1912), no sólo derrumbó las hipótesis
ameghinianas, sino que condujo al abandono de la búsqueda
sistemática de rastros de los primeros americanos en
las pampas argentinas junto con Ameghino, toda una línea
de investigación fue condenada al descrédito. Los
enigmáticos hallazgos de bolas de boleadoras y otros
artefactos aparentemente muy antiguos, que entre 1910 y
1930 efectuó el aficionado Lorenzo Parodi, en las
barrancas de Miramar (partido de General Alvarado,
provincia de Buenos Aires), fueron sospechados de fraude
y con ellos se fue diluyendo la idea de un poblamiento
muy remoto de la región. Sobre la base de múltiples métodos
de datación del pasado con que ahora se cuenta, la
cronología actualmente propuesta para el poblamiento de
América es bastante distinta de la que hace casi un
siglo suponía Ameghino. |
Hoy, una gran cantidad de
hallazgos arqueológicos apoya la hipótesis de que el
hombre llegó a este continente, cruzando el estrecho de Behring, entre Siberia y Alaska, hace 20 o 30 mil ańos.
Incluso los recientes hallazgos efectuados por Tom
Dillehay, de la Universidad de Kentucky, en el sitio de
Monte Verde (Chile) y por Niede Guidón, de l'Ecole de
Hautes Etudes en Sciences Sociales, en el estado de Piauí
(Brasil), han dado fechados que remontarían el
poblamiento de América del Sur hasta casi 35 mil ańos
atrás. Sin embargo, y a pesar de sus exageraciones
cronológicas, varias de las hipótesis de Ameghino han
vuelto a considerarse seriamente. Por un lado, el
descubrimiento en 1927 de puntas de proyectil clavadas
en huesos de bisontes extinguidos en Folsom (Nuevo México,
EE.UU.), llevó a que se aceptara que efectivamente el
hombre había coexistido con especies animales
extinguidas en América del Norte. Poco después, el
arqueólogo norteamericano Junuis Bird encontró en el
otro extremo del continente, en cuevas de la patagonia
chilena, otras puntas de proyectil, asociadas con restos
de caballo americano y de milodonte, ambos también
extinguidos. Por otra parte, algunas de las mismas
evidencias presentadas por Ameghino, como el cráneo
humano denominado por él como Diprothomo recuperado a
casi 15 m. de profundidad durante la construcción del
puerto de Buenos Aires, o el "esqueleto de
Fontezuelas" hallado en el interior de la caparazón
de un gliptodonte, no se explicaban dentro del modelo de
Hrdlicka de poblamiento de América en épocas
post-glaciales.
Las Extinciones
Pleistocénicas.
|
La
primera parte del período Cuaternario, el
Pleistoceno ha sido caracterizado como la época
de las glaciaciones, es decir por el avance de
grandes masas de hielo sobre los continentes
debido a un descenso de las temperaturas del
planeta. El comienzo de esta época ha sido
estimado entre dos y dos millones y medio de ańos,
mientras que su finalización llegaría hasta
hace diez mil ańos. Esta última fecha,
convencionalmente aceptada por los
investigadores de distintas disciplinas, marcaría
el fin de la última glaciación y el paso hacia
las condiciones climáticas más benignas que
caracterizan al período posterior en el cual aún
nos encontramos: el Holoceno o Reciente. Durante
el Pleistoceno se produce en África el origen y
la dispersión del género humano, como
resultado de un proceso de hominización que
habría comenzado algunos millones de ańos antes.
Esta época tuvo además una fauna característica,
cuyos representantes más conspicuos se
extinguieron abruptamente en América en el lapso
que media entre los 12.000 y 8.000 ańos antes
del presente.
En América del Sur, la fauna del Pleistoceno
incluía algunos representantes nativos,
aquellos que se habían originado y desarrollado
por decenas de millones de ańos en el
subcontinente, junto a otros "recién
llegados" provenientes de América
del Norte, cuando en la época anterior, el
Plioceno, se formó el istmo de Panamá.
|
 |
Entre
los primeros se encuentran los perezosos
terrestres como el megaterio, el milodon y el
glosoterio, los gliptodontes, lejanos parientes
de los armadillos actuales, el toxodonte, un
gran mamífero parecido al hipopótamo y la
macrauquenia. Entre el segundo grupo se destacan
tanto especies extinguidas (caballos americanos,
mastodontes y tigres "dientes de
sable") como vivientes (zorros, pumas,
guanaco, venados y ciervos). Gran parte de esta
fauna desapareció hacia fines del Pleistoceno
debido probablemente a los drásticos cambios
climáticos que se produjeron durante esta época.
|
Sin embargo, en América del Norte la presencia
recurrente de huesos de mamut y bisonte extinto
en los sitios arqueológicos más antiguos llevó
a pensar que la presión de caza de los primeros
pobladores americanos podría haber causado,
junto a otros factores, la extinción de estas
especies. En América del Sur, varios sitios
presentan también huesos de caballos
americanos, lo que plantea la posibilidad de que
el hombre también haya contribuido de alguna
manera a la desaparición de estas especies. ańos
en el subcontinente, junto a otros "reciénllegados"
provenientes de América del Norte, cuando en la
época anterior, el Plioceno, se formó el istmo
de Panamá. Entre los primeros se encuentran los
perezosos terrestres como el megaterio, el
milodon y el glosoterio, los gliptodontes,
lejanos parientes de los armadillos actuales, el
toxodonte, un gran mamífero parecido al hipopótamo
y la macrauquenia. Entre el segundo grupo se
destacan tanto especies extinguidas (caballos
americanos, mastodontes y tigres "dientes
de sable") como vivientes (zorros, pumas,
guanaco, venados y ciervos). Gran parte de esta
fauna desapareció hacia fines del Pleistoceno
debido probablemente a los drásticos cambios
climáticos que se produjeron durante esta época.
Sin embargo, en América del Norte la presencia
recurrente de huesos de mamut y bisonte extinto
en los sitios arqueológicos más antiguos llevó
a pensar que la presión de caza de los primeros
pobladores americanos podría haber causado,
junto a otros factores, la extinción de estas
especies. En América del Sur, varios sitios
presentan también huesos de caballos
americanos, lo que plantea la posibilidad de que
el hombre también haya contribuido de alguna
manera a la desaparición de estas especies.
|
Las
Huellas del Pasado.
Por: Cristina Bayon,
Universidad Nacional del Sur y Gustavo Politis,
Universidad Nacional del Centro de la Provincia de
Buenos Aires. Fragmento de la publicación original de
la revista de la Asociación Ciencia Hoy, Volumen 8 N°
48.
Desde el comienzo de las
investigaciones arqueológicas, se ha procurado
establecer la antigüedad y las características de las
ocupaciones indígenas en la costa de la región
pampeana. El tema ha tenido un desarrollo complejo, con
grandes controversias e intensos debates, sobre todo a
principios del siglo, cuando Florentino Ameghino postuló
una antigüedad geológica terciaria para los materiales
encontrados en los sitios costeros. Las explicaciones
han variado según las posturas teóricas de los
diferentes investigadores; para algunos arqueólogos,
como Osvaldo Menghin y Marcelo Bórmida, los yacimientos
se formaron por la presencia de los indígenas que vivían
y explotaban los recursos del litoral; por el contrario,
William Holmes y Francisco de Aparicio creían que los
sitios eran la evidencia de las periódicas visitas a la
costa atlántica de los indígenas que habitaban la
llanura pampeana. Sin embargo todos debieron hacer
frente a una misma dificultad: en el litoral bonaerense
los restos arqueológicos se encuentran sobre la
superficie del terreno. Por lo general, los materiales
aparecen apoyados en la arena de las hoyadas que forman
los médanos o sobre los afloramientos de sedimentos más
compactados. Hay dos casos excepcionales de depósitos
arqueológicos estratificados: por un lado, están los
controvertidos descubrimientos realizados en la década
de los veinte por Lorenzo Parodi, que nunca han sido
convalidados por los científicos, y por otro, las
sepulturas del llamado "Túmulo de Malacara",
que no han podido ser correlacionadas con los abundantes
materiales de superficie. Estos son, en realidad, lo
poco que ha llegado hasta nosotros de conjuntos mucho más
ricos y variados que, de haber estado protegidos bajo la
superficie del terreno, no hubieran sufrido el
importante deterioro producido por la acción de los
agentes naturales: viento, lluvia o sol. Por ejemplo,
los materiales orgánicos no se conservan a la
intemperie; la piedra y la cerámica, al quedar
expuestas, se mezclan permanentemente con los restos de
ocupaciones posteriores; en el caso de la alfarería, a
pesar de ser un material relativamente resistente, la
erosión termina por desintegrarla. Tampoco es posible
obtener fechas absolutas mediante la aplicación del
carbono catorce pues han desaparecido materias orgánicas
com el hueso, carbón vegetal o restos de plantas.
La situación cambió notablemente
en 1984 cuando se descubrió el sitio arqueológico La
Olla 1 y, posteriormente, otros cercanos y estrechamente
relacionados: La Olla 2 y Monte Hermoso 1. El conjunto
ha revelado datos sumamente novedosos para la región,
pues se han podido documentar centenares de pisadas
humanas, de aves y mamíferos, recuperar restos óseos
de varias especies de animales, artefactos de piedra y,
además, restos vegetales en un estado excepcional de
conservación. Los tres yacimientos mencionados están a
seis kilómetros al Oeste de la localidad de Monte
Hermoso (provincia de Buenos Aires), donde los depósitos
del fondo de una laguna de la época holocénica (unos
7.000 ańos antes del presente) aflorán en la playa
actual y dos veces al dia los cubre la marea. Se trata
de sedimentos estratificados de láminas intercaladas de
arcilla y arena, que se extienden unos 1.100m a lo largo
de la playa (figura 1); en el pasado formaban parte del
borde de una antigua laguna litoral, hoy desaparecida, y
que durante su proceso de formación fue visitada en
varias oportunidades por indígenas
cazadores-recolectores pampeanos que acamparon cerca de
sus márgenes.
żQué se ha conservado y cómo?
Los restos materiales preservados
en estos sitios son muy variados. En La Olla 1 y 2 se
han recuperado cientos de huesos de lobo marino, guanaco
y venado, valvas, caracoles marinos, fragmentos de
huevos de ńandú, restos vegetales, artefactos de
piedra y un objeto de madera decorado (figura 2).
Vicente Di Martino, director del Museo Municipal de
Monte Hermoso, descubrió en diciembre de 1983 el
yacimiento denominado La Olla 1. Por lo general el sitio
está tapado por la arena de la playa y excepcionalmente
queda expuesto; en esas ocasiones los materiales arqueológicos
se hacen visibles, pues afloran en los sedimentos
lagunares más compactados. Cuando la arena lo vuelve a
cubrir, es muy difícil excavarlo de manera sistemática:
el oleaje continuo y la arena suelta imposibilitan
mantener el área destapada. Di Martino y Luis Guzmán,
entonces director del Museo "José A. Mulazzi"
de Tres Arroyos, pudieron concretar a comienzos de 1984
una excavación rápida. Hubo que esperar hasta 1993
para que el sitio quedara expuesto y hacer una nueva
recolección y un estudio más detallado de los perfiles
geológicos. En el verano del 1995 los movimientos de
arena del litoral dejaron al descubierto un yacimiento
de características similares (La Olla 2), pero su
exposición fue tan breve que impidió cualquier tipo de
trabajo sistemático: sólo se recuperaron algunas
piezas óseas de lobo marino y unos pocos artefactos. En
ambos sitios de La Olla los huesos de lobo marino
pertenecen a las especies de uno y de dos pelos, y hay
una alta proporción de restos de los miembros
anteriores de los animales, especialmente húmeros y
radiocúbitos, mientras que son escasas las partes del
esqueleto axial. Esto sugiere que los lobos marinos
fueron despostados en el lugar donde los cazaron,
probablemente a la orilla del mar, y sólo el cuero y
algunos pedazos de carne fueron llevados para preparar y
consumir a la orilla de la laguna.
Para confeccionar los instrumentos
de piedra se usaron, principalmente, rodados costeros y,
en menor cantidad, cuarcita y arenisca. En algunos casos
por talla se redujo la roca a un tamańo y forma
convenientes, mientras que otros fueron trabajados por
desgaste o pulido, e incluso algunos tomaron forma por
el uso. El hallazgo más notable es un instrumento de
madera con decoración: se trata de una especie de espátula
larga adornada con líneas rectas y en zig-zag incisas.
Este espécimen, junto con dos estacas o puntas, son los
únicos artefactos arqueológicos de madera conocidos
procedentes de sitios pampeanos, y hasta ahora no hay en
el Cono Sur registro de objetos similares preservados
por el agua del mar.
En otro sector de la orilla de la
laguna, en el sitio Monte Hermoso 1, se ha conservado
una notable evidencia de la presencia humana. Se trata
de centenares de pisadas humanas (figura 3), de aves
(figura 5) y mamíferos, junto con restos de plantas,
semillas, algunas estacas o puntas de madera, caracoles,
cáscaras de huevo de ńandú y unos pocos huesos de
lobo marino. El sitio fue descubierto en 1990 por el geólogo
Rodolfo González, quien reconoció las pisadas humanas
en los sedimentos consolidados que afloraban a lo largo
de 800 metros en la playa actual. Las huellas son
claramente visibles en cuatro sectores principales y,
por ejemplo, en un área de 420m2 hay
impresas más de 400 improntas humanas, 20 de aves y dos
de artiodáctilos .
Además, las pisadas quedaron
impresas en por lo menos tres superficies antiguas
diferentes, es decir que la orilla de la laguna fue
transitada por los indígenas pampeanos en tres momentos
distintos.
żCuándo fue?
La excelente conservación del
material orgánico en los sitios ha permitido realizar
varios análisis de carbono catorce y, en consecuencia,
conocer cuándo ocurrieron los distintos eventos que han
quedado registrados en la laguna. Hasta el momento
disponemos de siete fechados radiocarbónicos que ubican
los acontecimientos entre 6.640 y 7.400 ańos antes del
presente, es decir a mediados del período Holoceno.
Para el sitio de La Olla 1 se hicieron dos análisis
sobre colágeno (proteína ósea) de fémur de lobo
marino, y dio una edad de 7.315 ± 55 ańos antes del
presente (AA-7972) y 6.640 ± ańos antes del presente (LP-303).
En La Olla 2 se utilizó una vértebra también de lobo
marino, y el resultado fue de 7.400 ± 95 ańos antes
del presente (AA-19292). Existen cuatro dataciones
radiocarbónicas obtenidas de muestras asociadas con las
pisadas en el sitio Monte Hermoso 1; una es una semilla
de una planta acuática (Ruppia sp.), extraída de las
fases arcillosas inferiores y dio una edad de 7.125 ±
75 ańos antes del presente (AA-7974). De la segunda,
hecha sobre una escápula de lobo marino incluida en los
bancos de areniscas, se obtuvo una antigúedad de 7.030
± 100 ańos antes del presente (LP-271). El tercero y
cuarto análisis proceden de los niveles estratigráficamente
más altos, se hicieron sobre un fragmento de rama de
una especie indeterminada y de unas semillas de Ruppia
sp. Las fechas obtenidas fueron 6.795 ± 120 ańos antes
del presente (AA-8699) para el fragmento de rama, y
6.705 ± 80 ańos antes del presente (AA-8700) para las
semillas.
żCómo era la laguna hace 7.000 ańos?
Carlos Zavala fue el responsable de
los estudios geológicos que se hicieron para determinar
el ambiente, el modo de depositación y la evolución
geológica de la laguna. Para esto se definieron y
estudiaron los cambios en el aspecto de los depósitos
mediante el análisis de las características internas
de los cuerpos de roca: estructuras sedimentarias
primarias, textura, contactos, color, geometría y
contenido paleontológico. Las características cambian
a lo largo del perfil, tanto vertical como lateralmente,
debido no sólo a las transformaciones ocurridas durante
la evolución de la charca y la formación del depósito,
sino también a las ligeras diferencias del ambiente en
que tuvo lugar la depositación.
|

Pisada humana en
el yacimiento paleoicnologico de Pehun-co en la
Prov. Buenos Aires. |

Recorrido de
enormes pisadas de Megatherium en el mismo
yacimiento único en el mundo. |
Los sedimentos que contienen las
pisadas están apoyados sobre depósitos de edad
pleistocénica -hace más de 10.000 ańos- expuestos en
la zona intermareal de la playa. La unidad que
corresponde al Holoceno se inicia con capas
esencialmente arcillosas de color gris claro, laminadas,
con delgadas intercalaciones de arenas y restos de
peces, roedores y vegetales. Aquí es donde se observa
la mayor densidad de pisadas. Por encima se disponen
sucesivos bancos de arena, entre los que se intercalan
delgadas láminas de arcillas. Es en estos contactos
donde se han detectado las pisadas mejor preservadas,
que son poco profundas y muchas conservan aún sus
rellenos.
La reconstrucción ambiental estuvo
a cargo de Silvia Grill y Dina Martínez, investigadoras
de la Universidad Nacional del Sur, a través del
estudio del polen y de organismos microscópicos acuáticos
(ostrácodos). Los análisis sugieren la presencia de
una vegetación de dunas costeras y cuerpos lagunares
tranquilos de interduna, con abundante material
nutritivo y buena oxigenación, que periódicamente sufría
episodios de inundación y de retracción. A pesar de
situarse muy cerca de la costa, la laguna
excepcionalmente tuvo comunicación con el mar. Si bien
se ha identificado paleomicroplancton marino
(dinoflagelados y acritarcas) en los sedimentos, su
pobre estado de conservación sugiere que se trata de
material redepositado. Los resultados del estudio del
polen seńalan que la flora del lugar se caracterizaba
por una vegetación herbácea de médanos que coexiste
con plantas acuáticas típicas de las depresiones
intermedanosas.
La identificación de cuatro taxa
de ostrácodos no marinos (Sarsc ypridosis aculeata,
Lymnocythere sp1, sp2, Cyprinotus salinus), indican que
la laguna era continental aunque salobre. El estado de
conservación, el número de especies y la asociación
faunística estaría evidenciando un cuerpo de aguas
tranquilas. En tanto que la precipitación de carbonatos
dentro de las valvas en algunas etapas, es un indicador
de importantes episodios de desecamiento y de un
ambiente inestable. La estratigrafía indica que además
de estas pulsaciones periódicas, la laguna era más
grande al principio, y que a lo largo de los siglos
paulatinamente se fue reduciendo. Hay indicios de que
hace unos 6.500 ańos la laguna era apenas un cuerpo de
agua somero de interduna y que estaba próxima a
desaparecer.
żPor qué se conservaron?
Para que se conserve un registro
arqueológico tan frágil como el de las pisadas
humanas, deben darse una serie de eventos en un orden
adecuado. En el caso estudiado, un ambiente de relativa
quietud (de baja energía), como las aguas de una laguna
costera, favoreció la preservación
El orden de los eventos fue el
siguiente:
1.- Debajo del agua se depositó
una capa de grano fino que fue importante para
registrar los detalles con mayor fidelidad.
2.- El sedimento de las márgenes,
al quedar expuesto al aire, perdió parte del agua de
los poros y adquirió cierta plasticidad, deformándose
parcialmente con el peso de los caminantes.
3.- Luego de impresas las
pisadas, el agua debió cubrirlas rápidamente pero
con baja energía. Las huellas que se conservaron no
quedaron expuestas por mucho tiempo, pues en caso
contrario se hubieran borrado. De este modo,
perduraron las más cercanas al borde de la laguna, y
es lo que determina que los sitios con pisadas estén,
casi siempre, en las márgenes de antiguos cuerpos de
agua.
4.- Con posterioridad a su
enterramiento, el depósito no sufrió grandes
transformaciones hasta que fue erosionado por un
agente "sutil" como el agua. En este caso,
el mar al penetrar entre la huella y su relleno hizo
que se separaran dejando a la vista la impronta. Sin
embargo, en la medida en que la erosión marina actúa
sobre los sedimentos con pisadas, las destruye. Al
principio remueve el relleno y la marca del pie queda
a la vista; luego, como la acción abrasiva continúa,
comienza a afectar la huella hasta que la borra
totalmente. Este proceso es casi imposible de detener,
pero en la medida en que el mar va erosionando algunas
pisadas, deja otras tantas al descubierto.
żDe quién son las huellas?
Se han hecho distintos trabajos
para conocer cómo se distribuyen en el sitio las
improntas y los restos materiales. Las tareas de
investigación se iniciaron en el extremo oriental,
porque es el más expuesto a la destrucción. Para un
estudio de este tipo, el terreno fue cuadriculado en áreas
de 1 metro de lado y luego, tomando el reticulado como
referencia, se midieron todas las pisadas. Hubo que
tener en cuenta varios datos: si se trataba del pie
izquierdo o derecho, el largo, el ancho, la profundidad
y la orientación de cada huella. Cuando se reconocía
un rastro, es decir una serie de huellas producidas por
la marcha de una misma persona, se tomó también la
distancia entre los pasos. Sobre la base de estos datos
se mapearon todas las pisadas, aisladas o formando
rastros . Con el objeto de hacer un análisis estadístico
que permitiera inferir los grupos de edad que estaban
representados, E. Navarro seleccionó un conjunto de
huellas que se estudiaron utilizando el análisis
multivaríado de componentes principales. El análisis
quedó restringido a aquellos casos que estábamos
seguros de que eran relativamente sincrónicos. El
sector elegido fue el área que estaba cruzada por el
rastro más extenso y todos estaban impresos en una única
superficie. De las huellas, cinco son de pisadas pequeńas
(14 a 17cm de largo) que seguramente corresponden a nińos;
otras cinco están formadas por improntas entre 19 y
23cm, que podrían corresponder a jóvenes o mujeres; sólo
una pertenece a un adulto de cierta estatura.
Otra característica importante es
que ninguna muestra una dirección determinada, ni son
paralelas. Esto indica que el sector de la orilla no fue
una senda, sino más bien un área por donde la gente
recorría una superficie amplia, no un camino lineal y
estrecho.
Imágenes del pasado
La interpretación propuesta a
partir de los datos actualmente conocidos, es que hace
siete mil ańos esta laguna tranquila de agua salobre,
cercana al mar, fue visitada periódicamente por indígenas
cazadores-recolectores de la región pampeana. El
paisaje costero formaba parte de su territorio y era
frecuentemente visitado ya que en las cercanías se podían
cazar lobos marinos, recoger rocas para confeccionar
instrumentos y, quizás, también recolectar conchas de
caracoles para hacer adornos. En distintas partes de la
antigua laguna han quedado las evidencias de las
diferentes actividades llevadas a cabo. La Olla 1 y 2
muestran los lugares donde se terminaban de procesar los
cuerpos de los lobos marinos. Las presas eran capturadas
en la costa marítima cercana y acarreadas ya
despostadas hasta la laguna. Allí se terminaba con el
trozamiento y, en parte, se consumían. La selección de
las porciones esqueletarias, las marcas de corte sobre
los huesos y los mismos huesos quemados testimonian
estas actividades. Es probable que también se
procesaran vegetales, lo que se infiere del tipo de
instrumental descartado en el lugar: rocas con caras
alisadas y pulidas usadas para moler o machacar semillas
y frutos. Las pisadas humanas de Monte Hermoso 1
muestran que las orillas de la laguna eran recorridas
por nińos, jóvenes y, tal vez, mujeres que deambulaban
en forma tranquila, sin una dirección definida. El
hecho de que sólo este segmento de la población esté
representado, es consistente con una actividad de
recolección en las márgenes de la laguna para
proveerse de los típicos recursos lacustres: plantas,
aves, huevos y peces. Entre los cazadores-recolectores
esa actividad la desempeńan, precisamente, mujeres y nińos.
Hasta ahora no se ha podido determinar el emplazamiento
del campamento, aunque la gran cantidad de huellas de nińo
indica que este debía estar próximo pues en esas
sociedades los ni-ńos no suelen alejarse solos a
grandes distancias de la vivienda. Estas evidencias
arqueológicas de la costa atlántica han permitido
reconstruir algunos aspectos poco conocidos de las
antiguas sociedades indígenas de la región pampeana.
Sin embargo aún no han aclarado un problema que desde
principios de siglo es largamente debatido: żestaban
estas poblaciones adaptadas a un único ambiente o, por
el contrario, la costa constituía un componente dentro
de un amplio circuito de nomadismo?
Sitios en peligro
Los yacimientos de Monte Hermoso y
La Olla se encuentran en peligro de destrucción porque
son sumamente frágiles frente a la acción del mar y de
la gente. La erosión marina es constante pues los
afloramientos se ven afectados diariamente por la marea,
y es aún mayor durante las frecuentes sudestadas. Por
el lado de la acción humana, el tránsito de vehículos
es el principal responsable de la destrucción, ya que
permanentemente circulan por el lugar desde tractores
hasta triciclones, y todos ellos de fuerte tracción; así,
las láminas que forman el depósito se resquebrajan y
el agua encuentra material fácil para erosionar. El mar
y los seres humanos se potencian en la destrucción
Otros sitios con huellas fósiles.
Desde los comienzos del siglo XIX se
conocen en la literatura científica sitios de distintos
períodos geológicos con huellas de vertebrados. Sin
embargo, las pisadas humanas son poco frecuentes. Las más
famosas y, probablemente, más antiguas son las de
Laetoli, en Tanzania, al este del África, que
corresponden a homínidos que vivieron hace unos 4 a 5
millones de ańos.
En América hay también huellas
humanas pero mucho más modernas; están, por ejemplo,
las de Acahualinca, Nicaragua, que fueron dejadas por
cazadores-recolectores a comienzos del Holoceno (unos
6.000 ańos antes del presente) en la orilla del lago de
Managua. Una gruesa capa de cenizas volcánicas de
varios metros de espesor cubrió las pisadas y permitió
entonces una excelente preservación.
Otra impronta famosa es la dejada
por un nińo en el piso arcilloso del sitio de Monte
Verde en el sur de Chile. Tom Dillehay, arqueólogo de
la Universidad de Kentucky (EE.UU.), estima que la antigüedad
de esta huella se corresponde con la de la ocupación
del sitio: aproximadamente 12.500 ańos antes del
presente. También se han registrado huellas y rastros
humanos en la zona de mareas de las costas de
Inglaterra, y cuya preservación se debe a procesos
similares a los de Monte Hermoso. En Formby Point,
estuario de Mersey al noroeste de Inglaterra, se
registraron pisadas aisladas y rastros que correponden a
dos períodos distintos: unas al neolítico (c. 4.000
a.C.) y otras a la edad del bronce (c. 1.500 a.C.).
Asimismo, en el sudeste inglés se han hallado huellas
humanas en el estuario de Severn que corresponden al período
mesolítico (c. 6.000 a.C.), junto con otras de ganado
vacuno: un animal que en los milenios posteriores
adquiriría vital importancia para los seres humanos.
En la Argentina, por su parte, la
costa del suroeste bonaerense posee un abundante
registro de pisadas antiguas y en Pehuencó se ha
descubierto el yacimiento paleoicnológico
[paleo=antiguo; icno= traza o huella; logos=ciencia] más
importante. Este sitio ha sido investigado por S.
Aramayo y T. Manera, quienes han registrado centenares
de pisadas de mamíferos y aves del Pleistoceno Superior
(12.000+/-110 ańos antes del presente) a lo largo de más
de 2km de afloramientos. La fauna representada es
notablemente variada y las pisadas son atribuibles, por
una parte, a animales que se han extinguido: megaterios,
milodontes, gliptodontes, camélidos de gran talla y
proboscidios; por otra, a especies actuales como cérvidos,
pumas, guanacos, un carnívoro semejante al aguará guazú,
ńandúes, flamencos, perdices y aves acuáticas
Lecturas sugeridas
BAYON, C. Y POLITIS, G., 1996,
"Estado actual de las investigaciones en el sitio
Monte Hermoso 1 (Prov. de Buenos Aires)", en
Arqueología (Instituto de Ciencias Antropológicas,
Facultad de Filosofia y Letras, Universidad de Buenos
Aires), 6:83-116.
POLITIS, G., 1989, "żQuién
mató al megaterio ?", en Ciencia Hoy; 1,2: 26-35
|
|