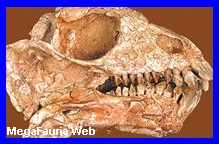
|
PaleoGuia del
Periodo
Paleozoico:
La era Paleozoica
abarca un lapso de 600 a 245 millones de ańos antes del presente. A
diferencia a las Eras posteriores, la Era Paleozoica no la hemos
dividido por Periodos o Épocas, para simplificar, aunque
posiblemente mas adelante lo aremos. Usted puede colaborar con esta
sección, escribanos a
grupopaleo@gmail.com
|
|
Durante la primera
parte de la Era Paleozoica (Cambrico, Ordovícico y Silurico), el
continente sudamericano estaba unida a África, Antartida, Australia
e India formando la masa continental de Gondwana que estaba separada
por el mar del norte América, Europa y Asia, que constituían
bloques continentales independientes, ubicados en la franja
ecuatorial. En este momento la Republica Argentina probablemente
haya estado cubierta casi por completo por las aguas. En el
Devonico, America del norte y Europa se unieron en un único bloque
llamado Euroamérica e iniciaron una lenta deriva hacia Gondwana,
con la cual comenzó a funcionarse en el Carbonífero. Gondwana en
ese momento se encontraba ubicada sobre el polo de rotación con lo
cual una gran parte de las tierras estaban cubiertas por gruesos
mantos de hielo. Durante el Carbonífero las condiciones climáticas
eran muy cálidas y húmedas. El sustrato estaba permanentemente
anegado y abundaban los pantanos. En el periodo Permico, Euroamerica
se fusiono finalmente con Gondwana, formando el supercontinente
Pangea. Recién a fines de este periodo, los anfibios y reptiles que
se habían originado en Euroamerica en el carbonífero comenzaron a
dispersarse hacia Gondwana.
<<<
Principales sitios
fosilíferos de Argentina.
|
|
Si bien la mayoría de los
organismos experimentaron grandes radiaciones adaptativas durante este periodo,
hacia fines del mismo se produce la gran primera extinción
masiva. La vida marina experimento una crisis con la extinción gradual de
Trilobites, Fitoplancton, Braquiópodos, Nautiloideos, Briozoos,
Cnidarios y Equidermos. Sin embargo en los continentes las
condiciones climáticas eran favorables para el desarrollo de vida
mas compleja.
|

Apankura machu.
Vaccarl,
Edgecombe & Escudero,
2004.
|
 |
|
 |
|
Fósil de
Apankura machu
hallado en el Cambrico de Jujuy. Imagen de prensa.
|
|
Reconstrucción de
Apankura machu
(animada en YouTube)
realizada por Carlos E. Gómez. |
Cambrico. Los restos fósiles de
un artrópodo -un ancestro mandibulado de los cangrejos, las
langostas marinas, los ciempiés y los insectos- que vivió hace unos
495 millones de ańos, fueron descubiertos en Jujuy por
investigadores argentinos del Conicet, quienes publicaron sus
conclusiones en un estudio publicado por la revista Nature. Se trata
de un artrópodo fósil hallado en la localidad de Tilcara, en la
Quebrada de Humahuaca, al que denominamos Apankura machu,
que significa cangrejo viejo en quechua. Apankura fue
hallado en la Garganta del Diablo por Cristian Escudero, un
estudiante de geología de la localidad de Palpalá (en Jujuy) que
luego lo llevó al laboratorio para su análisis. Otro aspecto muy
importante de esta investigación fue el hallazgo de una traza fósil,
probablemente realizada por Apankura. Estas trazas
-destacó el científico- son las huellas o marcas del desplazamiento
de los organismos dejadas en el sedimento. Lo llamativo es que se
trata de huellas similares y de la misma antigüedad que las
encontraron petrificadas en rocas que se formaron a partir de dunas
de arena en una antigua playa cerca de lo que hoy es la provincia de
Ontario, en Canadá.
Archaeocyatha.
Vologdin, 1937.
 |
 |
 |
|
En este caso,
Archaeoscyphia sp.una esponja del Ordovícico
medio, hallado en la precordillera en San Juan. |
Fosiles in situ de Archaeocyatha.
Prensa. |
Aspecto de Archaeocyatha
(*). |
Cámbrico. Fueron
animales pequeńos, de varios centímetros, y que poseían
forma cónica o cilindro-cónica. Su posición sistemática es
incierta, habiéndose incluido con los poríferos (filo
Porifera) o en un filo propio (filo Archaeocyatha).
Exclusivos del Cámbrico inferior (570 a 536 Ma). Junto a
algas calcáreas, fueron formadores de arrecifes en ambientes
marinos someros. Distribución geográfica muy extensa:
aparecen fósiles de arqueociatos en todos los continentes.
Poseen una capa o muralla externa y en la mayoría de las
familias también una muralla interna. El espacio entre las
dos murallas se denomina intervalo, dividido por tabiques
radiales denominados septos. No se parecen directamente a
las esclerosponjas. La disposición de los poros consiste en
un diámetro menor en la muralla externa que en la interna.
De esta manera, el agua circulaba por los poros de fuera
hacia el interior. Este patrón es típico de las esponjas.
Así se aceptó la cercanía entre arqueociatos y
esclerosponjas. Hoy se considera un grupo extrańo. A la
altura del río Colorado, precisamente, hay un arco
volcánico que atraviesa la Patagonia y pertenece al
Paleozoico superior, se han encontrado interesantes
fósiles de Archaeocyatha). En efecto, en
la zona de El Jagüelito, una localidad en la provincia de
Río Negro, cerca de Sierra Grande, el doctor Pablo González,
investigador del Conicet, encontró
restos fósiles de arqueociátidos, unas esponjas primitivas, de
forma cónica y unos pocos centímetros de longitud. Era la
primera vez que se encontraban, en América del Sur, estos
organismos que son exclusivos del período Cámbrico inferior.
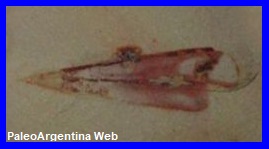
Hyolithes amosi.
Gonzalez & Sabattini, 1972.
|
 |
|
.jpg) |
|
Fósil de Hyolithes.
Ilustrativo (*) |
|
Aspecto del primitivo
Hyolithes (*) |
Cámbrico.
Los Hyolitha son una clase de pequeńos animales lofoforados
de concha con forma cónica que vivieron en la era Paleozoica. Sus
fósiles se encuentran desde el inicio del Cámbrico hasta el final
del Pérmico, aunque son poco abundantes a partir del Ordovícico.
Durante mucho tiempo fueron considerados moluscos o afines a ellos.
Las conchas calcáreas, probablemente aragoníticas, son cónicas, de
sección triangular o semicircular y normalmente aplanadas en la zona
ventral. Poseían una cubierta u opérculo, normalmente articulado. En
algunos fósiles de las Lutitas de Burgess, que conservan impresiones
de partes blandas, se ha podido observar que poseían un lofóforo
extensible con tentáculos, similares a los de las larvas de los
braquiópodos, usado como órgano de alimentación, y un estómago largo
en forma de U. Tenían hábitos solitarios, epibentónicos semi-sésiles,
es decir vivían sobre el sustrato del fondo marino con movilidad muy
reducida, y eran suspensívoros, se alimentaban de partículas en
suspensión en el agua que atrapaban con los tentáculos del lofóforo.
También se conoce Hyolithes patagoniensis. En estos
sedimentos se los encuentra asociados a
Saltarella
sp, o al braquiopodo
Billingsella, Algunos
fósiles de Hyolithes proceden del Paleozoico Superior
del Grupo Tepeul, Provincia del Chubut, y precordillera de Mendoza y
San Juan.
Chancelloria
eros. Walcott, 1920.
|
 |
|
 |
|
Fósil de
Chancelloria sp. (*) |
|
Aspecto de Chancelloria sp del Cambrico. |
Cámbrico. Es
un género de animales primitivos conocidos del Cámbrico. Fue
descrito por primera vez en 1920 por Charles Doolittle Walcott ,
quien los consideró como uno de los grupos más primitivos
de esponjas. Esto parece poco probable, y actualmente se encuentra
en el enigmático grupo Coeloscleritophora. Muchos de estos fósiles
se componen únicamente de espinas y otros fragmentos, y no es seguro
que pertenezcan al mismo tipo de organismo. Otros especímenes
parecen ser más completos y representar organismos sésiles en forma
de bolsa con una piel suave blindada con escleritos calcáreos
en forma de estrella de los cuales irradian espinas afiladas.
Algunos paleontólogos los clasifican como esponjas, una idea que el
estilo de vida sésil y la estructura simple de los canciloridos
hacen plausible. Otras propuestas sugieren que eran más avanzados, o
al menos se originaron en antepasados más avanzados, estando
relacionados con formas de babosas. Probablemente vivían en fondos
marinos fangosos, ya que sus escleritas aumentan de tamańo de abajo
hacia arriba, y todos tenían engrosamientos en las bases, que se
consideran anclas. Se registran en Quebrada Empozada en el área San
Isidro de la Precordillera Austral, al oeste de Mendoza, como así
también, en Cordillera Chica de Zonda, en la Precordillera Oriental
cerca de San Juan y en la Formación Los Sombreros en la Cordillera
Tontal de la Precordillera Occidental. Otras de las esponjas
conocidas para el Cambrico local es
Girvanella tubules.
Beltanelliformis sp.
Menner, 1974
|
 |
|
 |
|
Aspecto
de un fósil de
medusoides, del Cambrico de Villavicencio, Mendoza. |
|
Beltanelliformis, en vivo
(*). |
Cámbrico.
Es un género de
fósiles de medusoides. Son aquellos seres extintos que han sido
descritos como similares a medusas y que habitaron al final
del Precámbrico, durante el período Ediacárico y principios del Cámbrico.
Medían menos de un centímetro. Se considera que serían los animales
más antiguos, conjuntamente con los petalonamos, pues se han
encontrado fósiles desde hace aproximadamente 580 millones de ańos
de antigüedad. Inicialmente se pensó que fueron nadadores, en
realidad se concluyó posteriormente que eran como pólipos o discos
bentónicos anclados al fondo marino, o endobentónicos
semienterrados. Estos fósiles no presentan claramente
características típicas de las medusas como tentáculos bien
definidos, marcas musculares, gónadas, ni presencia de cnidoblastos.
Hoy se consideran un enigma evolutivo, no se les ha asignado un filo
y podrían estar relacionados con el hipotético grupo Vendobionta,
dada su antigüedad y simplicidad. Beltanelliformis,
proceden de afloramientos en la Cordillera Oriental de las
provincias de Tucumán, Jujuy y Salta, acompańados además de otros
icnogéneros como Nereites, Tasmanadia, Helminthoidichnus,
Taphrohelminthopsis y Glockerichnus.
Thysanopyge argentina.
Kayser 1898.
|
 |
|
 |
| Ejemplar de
Thysanopyge taurinus, de la localidad de Pascha, de la
provincia de Salta. |
|
Reconstrucción en vida del primitivo Trilobite Thysanopyge,
procedente de la Provincia de Salta, Republica Argentina. |
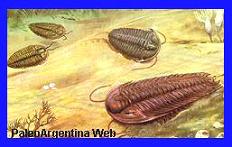
Cámbrico
-
Ordovícico.
Trilobites de gran tamańo que vivió durante el Ordovícico de la
Formación Acoste y Pascha en la Provincia de Salta. La antigüedad de
los presentes sedimentos corresponden a una antigüedad
aproximadamente los 450 millones de ańos. Como ya hemos citado la
descripción general del presente organismo, no desarrollaremos mas
información al respecto, aconsejando leer mas arriba. Algunas
especies bien registradas en el Cambriano Argentino son: Olenellus
sp, Mendospidella
digesta, Zacanthoides ferula, Chilometopus parabolicus, Asperocare
argentinum, Glossopleura, Prometeoraspis, Williamsinaornata y
Talbotinella communis.
Tetragraptus s.p.
Salter, 1863.
|
Graptolites.
Tratragraptus s.p del Ordovicico - Silurico (izq). Al
Lado, Caliza Estromatolitica del Precámbrico de la Provincia
de Buenos Aires. Ambos exhibidos en el Museo de La Plata.
|
| |
Ordovícico - Silurico. Graptolites. Son estructuras
estratificadas de formas diversas, formados por la captura y
fijación de partículas carbonatadas por parte de
cianobacterias en aguas someras que, en la fotosíntesis,
liberan oxígeno y retiran de la atmósfera grandes cantidades
de dióxido de carbono, que, en parte, emplean en la
construcción de los estromatolitos. El espesor de las
láminas es inferior a algunos milímetros y su forma es plana
(este tipo de estructura se denomina laminación algal o
criptalgal) a hemisférica y columnar. Suelen presentarse
numerosos poros entre las láminas (porosidad fenestral). Se
encuentran estromatolitos fósiles en todas las eras
geológicas y uno de los indicios más antiguos de vida en la
Tierra son estructuras estromatolíticas de hace 3500
millones de ańos encontradas en Warrawoona (Australia).
Los estromatolitos o camas de
piedra son fruto de células que se agrupan en colonias
formando rocas sedimentarias. Las células fosilizadas más
numerosas se encontraron en tales rocas formadas al borde de
mares cálidos. Al examinarlos en corte, al microscopio, se
distinguen muchas capas superpuestas en finas láminas
apiladas unas sobre otras en las que sólo la capa
superficial contiene organismos vivos. Estas rocas son pues
el resultado de la unión de minúsculos seres unicelulares,
cianobacterias, que viven en mares cálidos y en aguas poco
profundas. Las rocas se forman muy lentamente, capa sobre
capa, al morir las células de una capa, depositarse el
carbonato cálcico de sus paredes sobre la capa anterior y
formarse sobre ellas una nueva capa viva. Hace unos 3500
millones de ańos, cuando en los océanos ya emergían millones
de células vivas, aparecieron los estromatolitos. Ya hace
2500 a 1000 millones de ańos atrás, los arrecifes de
estromatolitos estaban ampliamente expandidos y comenzaron a
segregar un gas que fue causante de la primera extinción
masiva del planeta. Este gas era el oxígeno y provocó un
cambio drástico en la Tierra, notable hasta nuestros días.En
octubre 2009 la bióloga e investigadora del CONICET María
Eugenia Farías y su equipo descubrieron a 3000 m de altura
en Socompa y Tolar Grande, Salta estromatolitos.
Investigadores de la NASA luego redescubrieron
estromatolitos vivos en la zona de la Puna Salteńa, en el
norte de Argentina, más precisamente en la laguna de Socompa
y en seis "ojos de mar" (lagunas pequeńas, profundas y muy
saladas) cercanos a la población de Tolar Grande en el
departamento Los Andes. La particularidad de estos
estromatolitos es que el ambiente en el que se forman es
extremo, con alta radiación ultravioleta, muy salino y con
arsénico.
Araneograptus murrayi.
J. Hall, 1865.
|
 |
 |
 |
|
Araneograptus murrayi.
Ejemplar del Museo de Ciencias Naturales de Salta.
|
Aspecto de fósil de
Araneograptus
murrayi. |
Aspecto colonial de
Araneograptus
murrayi. |
Ordovícico.
Esta especie de graptolites integra el grupo
de los hemicordados. Son organismos coloniales y solitarios
que se originaron en el cámbrico inferior, hace 540
mill0ones de ańos y perduran hasta la actualidad. Lo que se
preserva en estado fósil es la estructura inorgánica o
colonia propiamente dicho, las cuales alojan a sus
habitantes los zooides.
Araneograptus
murrayi,
es una colonia constituida por ramas o estirpes que
presentas habitáculos para los zooides llamadas tecas. Eran
de hábitos planctónicos suspendidos por medio de un delgado
filamento denominado nema, que culmina en un disco de
flotación. Algunos materiales de
Araneograptus
murrayi,
provienen de
la Sierra de
Zenta, Cordillera Oriental y de la
región de la
Sierra de Cajas, Departamento Humahuaca en la Provincia de
Jujuy
y en Tarija, Bolivia. Otros géneros asociados;
Hunnegraptus copiosus Kiaerograptus, Adelograptus
y Paradelograptus antiquus.
Climacograptus bicornis.
J. Hall, 1847.
|
 |
|
 |
|
Roca
con fósiles de Climacograptus bicornis del
Ordovícico de de San Isidro, Las Heras y Cerro
Bola, San Rafael de Mendoza.
Museo de Ciencias
Naturales y Antropológicas Juan Cornelio
Moyano. |
|
Recreación de una colonia de Graptolites. (*). |
Ordovícico medio-superior.
Estos graptolites
de la familia diplograptidae son fósiles comunes y tienen
una distribución mundial. La preservación, cantidad y cambio
gradual permite que se usen como fósiles guía para datar los
estratos de rocas en todo el mundo. Durante el Paleozoico
los graptolites evolucionaron rápidamente y dieron lugar a
muchas especies diferentes. Los geólogos británicos pueden
dividir las rocas de los períodos Ordovícico y Silúrico en
biozonas de graptolitos; que por lo general tienen una
duración menor de un millón de ańos. Una glaciación en todo
el mundo al final del Ordovícico eliminó la mayoría de las
especies de graptolites que vivían entonces; las especies
presentes durante el período Silúrico fueron el resultado de
la diversificación de sólo una o dos especies que
sobrevivieron a la glaciación del Ordovícico. Los
graptolites también se utilizan para estimar la profundidad
del agua y la temperatura en la que vivían estos organismos.
Cada colonia de graptolites (conocida como rabdosoma) tiene
un número variable de ramas (llamadas estipes) originados
desde un individuo inicial (llamado sícula). Los individuos
subsiguientes (zooides) se alojan en una estructura tubular
o con forma de copa.
Orthidae.
Woodward, 1852.
|
 |
 |
|
Ejremplar exhibido en el Museo de Ciencias Naturales
"Miguel Ángel Arra" de la Universidad Nacional de Salta. |
Aspecto y forma de vida de los Ortidos. Museo de
Ciencias Naturales "Miguel Ángel Arra" de la Univ. Nacional de Salta. |
Ordovícico. Los
ortidos eran invertebrados marinos bentónicos sesiles
braquiópodos, que
vivieron en distintas profundidades. Fueron muy importantes
y abundante durante el Cambrico - Ordovícico. Tenían una
conchilla calcárea formadas por dos valvas de contorno
semicircular y se adherían al fondo marino por medio de un
pedúnculo muscular. En su parte externa suelen mostrar
ornamentaciones como costillas que divergen
radialmente desde el umbon. La roca que posee estos fósiles
cuya foto acompańa el presente texto, es portadora de
braquiópodos ortidos especialmente, y depende de depósitos
sedimentarios del Periodo Ordovícico superior, unos 445
millones de ańos aproximadamente, muy característicos del
noroeste argentino, y fueron hallados en Sierra de Mojotoro,
Provincia de Salta (Capital).
Lituites
s.p.
Bertrand, 1763.
|
.Lituites.
Este genero es conocido a partir de algunos ejemplares
recuperados en el río Chacritas en la provincia de San Juan,
Argentina. Imágenes ilustrativas. Recreación tomada de
vilaine.de. |
Ordovícico
- Silúrico. Fueron los
depredadores de mayor tamańo en los mares del ordovícico, hace 450
millones de ańos. Se encuentra en las aguas tropicales entre los océanos
Índico y Pacífico. A diferencia de otros moluscos cefalópodos,
como el pulpo o el calamar, el nautilo tiene unos ojos simples, sin
cristalino. También, en lugar de un par, tienen dos pares de
branquias y hasta 90 tentáculos dispuestos en dos círculos
alrededor de la boca. Pero por lo que más se caracterizan es por su
envoltura o concha lisa y enrollada, de hasta 28 cm de diámetro.
Está forrada con nácar y dividida en una serie de compartimentos
progresivamente mayores, el último de los cuales les sirve de
vivienda. Las paredes (septos) que forman las cámaras están
perforadas por un tubo (sifón) que está unido al animal. El
intercambio de líquidos y gases que se produce a través de sus
paredes, permite al nautilo regular la posición hidrostática.
Parece que el nautilo descansa durante el día en el fondo del océano
a unos 600 m de profundidad. Por la noche nada por los alrededores
forzando la salida de agua a través de una especie de embudo
primitivo que posee para este fin y se alimenta de diatomeas,
quisquillas y algas. Cuando se está alimentando mantiene sus tentáculos
extendidos para atrapar todo lo que se acerque a ellos. Los tentáculos
son pequeńos, contráctiles y adhesivos, pero no tienen ventosas.
Se conoce poco acerca del comportamiento y el ciclo de vida de este
molusco.
Maclurites sp. Le
Sueur, 1818.
|
 |
 |
|
Roca con conjunto de restos fósiles de Maclurites,
del Ordovícico de la precordillera de la Provincia de San
Juan.
Museo Paleontológico de la Universidad Nacional de Córdoba. |
Fósil de
Maclurites (*) |
Ordovícico.
Es un género extinto de los gasterópodos Ordovícicos
(caracoles) encontrados como fósiles y útiles para las correlaciones
estratigráficas (el Período Ordovícico duró desde hace
aproximadamente 488 millones a 444 millones de ańos). El caparazón
se enrolla distintivamente y se reconoce fácilmente. Las
Macluritas también tenían un opérculo, o segunda caparazón,
que cubría la abertura de la caparazón más grande del cuerpo.
Maclurites es característico de un grupo de gasterópodos
tempranos que apareció por primera vez en el Cámbrico Tardío y se
extinguió al final del Ordovícico.
Alunas muestras muy importantes fueron recuperadas en el Ordovícico
de la precordillera de la Provincia de San Juan.
Rusophycus
polonica.
Seilacher 1970
.
|
 |
 |
 |
|
Roca con
restos de reposo de Trilobites (Rusophycus)
en el Museo Paleontológico de
Córdoba. |
Rusophycus en roca cuarcitica en un espigon
costero. Museo de Miramar. |
Recreación de
la formación del icnofosil de
Rusophycus. |
Ordovícico.
Es un paragénero de icnofósiles presente en rocas
sedimentarias de facies marina profunda, marina superficial, fluvial
y lacustre. Es uno de los icnofósiles más representativos de la
actividad etológica de trilobites si bien son realizados por una
gran variedad de organismos. Las trazas de Rusophycus
son característicamente bilobuladas con simetría bilateral, de
pequeńo o mediano tamańo, de altura variable y que pueden ser
perfectamente paralelas entre sí o fusionarse en su zona posterior.
Los dos lóbulos pueden aparecer ornamentados con pequeńas
estriaciones trasversales y, más raramente, con estriaciones
longitudinales externas. Se corresponden con trazas de reposo (tipo
cubichnia) al funcionar como cubil o madriguera, de
alimentación (tipo fodinichnia) al ser marcas de
remoción de sustrato en busca de alimento o de depredación (tipo
praedichnia) al ser marcas realizadas al acechar un
organismo su presa. Algunos investigadores incluso han sugerido que
pudieran tratarse de nidos excavados para depositar en ellos
huevos. Estos icnofósiles fueron interpretados en un principio junto
a Cruziana como fósiles de vegetales. En 1864 John
William Dawson publicó un trabajo demostrando que eran rastros de la
actividad excavadoras de trilobites. Posteriormente en 1873 un
estudio similar fue dado a conocer por Alfréd Gabriel Nathorst con
más éxito que su predecesor. Ha sido registrado en el
Ordovícico temprano (480 millones de ańos) de la Cordillera Oriental, Puna, Sierras Subandinas, Precordillera, Ventania y Tandilia. Argentina.
Arthrophycus
alleghaniensis. Harían.
|
 |
 |
|
Roca
cuarcitica en un espigon costero con muestras de
Arthrophycus. Museo Punta Hermengo de Miramar. |
Detalles de
Arthrophycus alleghaniensis. |
Ordovícico.
Es un icnotaxón común en entornos
silícico-marinos Ordovícico-Silúrico de poca profundidad. Se
los considera madrigueras oblicuas a horizontales,
transversales y ramificadas. Las madrigueras son paralelas o
subparalelas, a veces con extremos afilados. Se puede encontrar
solitario, pocas madrigueras o como un grupo. Pudo servir como
refugio o
depósito de alimentación. Solo en ambientes de origen
marinos de poca profundidad. Se considera como el organismo
responsable de estas estructuras a anélidos o artrópodos. Se los
puede hallar desde Cámbrico al Carbonífero.
La presencia de Arthrophycus minimus en la Formación Santa Rosita del
noroeste de Argentina indica que Arthrophycus se extiende al menos
desde el Cámbrico Superior al Cámbrico inferior y, por lo tanto, su utilidad
bioestratigráfica se extiende. ichnospecies más jóvenes, Arthrophycus
minimus,
Arthrophycus brogniartii, Arthrophycus
alleghaniensis y Arthrophycus lateralis.
Mimetaster florestaensis.
Aris et al. 2017;
|
 |
|
 |
|
Fósil de
Mimetaster florestaensis. Foto de
Josefina Aris. |
|
Aspecto de
Mimetaster florestaensis. |
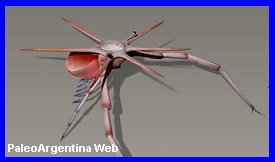
Ordovícico.
En la sierra de Mojotoro (provincia de Salta) fue hallado
recientemente el fósil de un animal perteneciente a un grupo de
artrópodos con un registro muy pobre, los marrellomorfos. Los restos
fósiles pertenecen a una nueva especie, bautizada con el nombre de Mimetaster
florestaensis, y corresponden al Tremadociano (Ordovícico) de
Argentina.. La especie salteńa está emparentada con otra del género Mimetaster que
procede de rocas devónicas de Alemania El hallazgo de Mimetaster
florestaensis es muy importante por varias razones. En primer lugar,
se trata de la primera cita de este tipo de artrópodos para
Argentina y Sudamérica, lo que completa el registro del grupo a
nivel mundial, ya que faltaba encontrarlo en Sudamérica; por otra
parte, a lo anterior se le suma el hecho que se trata de una nueva
especie, lo que incrementa el número de especies del grupo de los
marrellomorfos; además, se pudieron establecer relaciones de
parentesco de M. florestaensis con otras especies de otras partes de
mundo; y finalmente es importante seńalar que esas relaciones de
parentesco establecidas fueron nuevas evidencias paleobiogeográficas.
Los marrellomorfos son un grupo de artrópodos no trilobites marinos.
Se originaron en el Cámbrico (hace 541 millones de ańos) y el último
registro data del Devónico (383 millones de ańos). Su cuerpo estaba
aplanado en sentido dorso-ventral lo que significa que habitaban los
fondos marinos con probables incursiones nectónicas7. Tenían el
cuerpo segmentado con importantes escudos cefálicos de diferentes
tamańos y formas.
Palaeophycus vulgaris. Borrello, 1964.
| |
|
|
|
 |
|
 |
|
Mariano Magnussen Saffer, con trazas de
enormes anélidos marinos en rocas cuarciticas en Miramar. |
|
Aspecto de algunos
anélidos y otros invertebrados registrados en el Devonico (*). |
Ordovícico.
Se trata de un icnogenero o rastro primitivo. Si bien están catalogados como
otros icnofosiles paleozoicos de la provincia de Buenos Aires como "fósiles
problemáticos", es probable que estas marcas se deban a la pistas originadas por
la traslación de anélidos marinos de tamańo grande. Recientemente, Mariano
Magnussen del Museo de Ciencias Naturales Punta Hermengo hallo cerca del muelle de
pescadores de la ciudad de Miramar, una gran roca de cuarcita, depositada allí
artificialmente, con el propósito de construir la defensa costera, la cual
portaba una muestra del icnogenero Palaeophycus. Esta roca que se
emplea comúnmente en toda la costa bonaerense para este fin, proceden de las
canteras ubicadas en la localidad de Chapadmalal. Se caracterizan por sucos
semirellenos de sutil trazado, flexionados, superpuestos con sección regular
mostrando bordes netos, mostrando a veces giros y superposiciones sumamente
variada. Antecedentes de estos hallazgos corresponden a la Estancia La Celina en
la provincia de Buenos Aires en 1945. Se encuentran en sedimentos de arenas
cuartiticas de grano fino. El promedio de medida de estas trazas es de 1 metro
de longitud y entre1 y 2 centímetros de ancho. En oportunidades se encuentra
asociado a trazas de Corophioides y Tigillites.
Cochlioceras avus.
Eichwald, 1860.
 |
 |
 |
|
Conjunto de fósiles de
Belemnites
in-situ. Ilustrativo. |
|
Reconstrucción de un
Belemnite |
Ordovícico.
Los belemnites,
grupo extinguido de moluscos cefalópodos emparentados con la sepia.
Aunque en el carbonífero (hace unos 345 millones de ańos)
surgieron animales parecidos, los verdaderos belemnites no empezaron
a ser abundantes hasta el principio del jurásico (hace 195 millones
de ańos). Todos los belemnites se extinguieron al final del cretácico
(hace 65 millones de ańos). Vivían en el mar, agrupados en bancos
y alimentándose de otros animales suficientemente pequeńos para
poder ser capturados y dominados por ellos. El cuerpo del belemnites
tenía forma de torpedo rematado en una cabeza grande y con brazos
fuertes provistos de ganchos para sujetar a las presas. Como todos
los cefalópodos, los belemnites se mueven a reacción, emitiendo un
chorro de líquido. Al igual que el calamar y la jibia, podían
librarse de sus depredadores (tiburones e ictiosaurios) lanzando
tinta para ocultarse mientras huían a un escondite seguro; esto se
sabe porque algunos fósiles excepcionalmente bien conservados
tienen los restos del saco de tinta. Los belemnnites tenían una
concha dividida en cámaras y llena de gas, llamada fragmocono. Esto
les permitía flotar entre dos aguas, de forma parecida a los
ammonites, con los que compartieron los mares de la antigüedad.
Pero la concha de los belemnites se diferencia de la de los
ammonites por la estructura interna, totalmente recubierta de piel y
músculo. Además del fragmocono, la concha de los belemnites tenía
una estructura sólida de forma de bala llamada rostro, que solía
encerrar parte del fragmocono y estaba dispuesta en la parte
posterior del animal. Como estaba formado por un fragmento sólido
de calcita, el rostro se fosilizaba fácilmente y es ahora el resto
más común de belemnites. Antiguamente se llamaba ‘rayo del
diablo’, pues se creía que se formaba cuando un relámpago
alcanzaba el suelo. Se piensa que el rostro evolucionó como
contrapeso de la voluminosa cabeza y los brazos, que ocupan el
extremo opuesto del fragmocono. Además, las acanaladuras de los
lados del rostro pueden indicar que el belemnites tenía aletas,
quizá utilizadas como instrumento de elevación, dirección y
freno. Los rostros de los belemnites del jurásico tienen una forma
característica de cono o de cigarro. Durante el cretácico muchos
adoptaron una forma más parecida a una lágrima, como en
Actinocamax, común en las cretas de Gran Bretańa y el resto de
Europa. El belemnites del cretácico Duvalia tenía un rostro
aplanado por dos lados, parecido a una hoja de cuchillo. En
Argentina, sur de Mendoza, Neuquén, Chubut y Santa Cruz. Se han
recuperado fósiles de Belemnites Conchioceras avus,
proceden del ordovícico de
Sierra de Villicúm,
en la precordillera de la Provincia de San Juan. De la misma zona,
fue registrado mas tarde
otro
orthocerado,
Braulioceras sanjuanense (Kröger
et al. 2007), de unos 465 millones de
ańos.
Orthoceras sp.
Aparicio, 1969.
|
 |
|
 |
|
Ortocheras. Quebrada de
Talacasto, (San Juan). Colección del Museo de Paleontología
de la Universidad de Córdoba. |
|
Aspecto en
vida del extrańo
Ortocheras. |
Ordovícico.
Es
un género extinto de moluscos cefalópodos. Fueron muy abundantes en
los mares del Paleozoico. Los más antiguos datan del Ordovícico
superior, hace unos 450 millones de ańos, y se extinguieron a
finales del Triásico, hace unos 208 millones de ańos. No eran muy
diferentes a los actuales, nautilos aunque presentaban una concha
externa de forma cónica acabada en punta que estaba dividida en
cámaras. Podían llenar y vaciar de agua las cámaras internas, de tal
manera que podían situarse a distintas profundidades, dependiendo de
ese llenado. Este género se denomina a veces Orthoceratites, y a
veces mal denominado como Orthocera u Orthocerus. Probablemente fue
pariente del Cameroceras. Sus presas favoritas eran los Trilobites,
pues eran fáciles de atrapar y abundantes. Se ve que el
Ortocheras era muy voraz, pues se han encontrado tres
trilobites dentro de uno de estos cefalópodos. Se han recuperado
muestras de este animal en las Sierras de San Rafael, provincia de
Mendoza, Quebrada de Talacasto, provincia de San Juan, y en todo el
flanco oeste de la Precordillera Argentina.
Sacabambaspis janvieri. Gagnier
et al. 1986.
|
 |
 |
 |
|
Ejemplares de Sacabambaspis janvieri. del Ordovicico de V,
Bolivia. |
Ejemplar de
Sacabambaspis sp. |
Posible
aspecto del primitivo pez Sacabambaspis janvieri (*). |
Ordovícico.
Los peces son el grupo de
vertebrados más numerosos que existe, la enorme diversidad de
formas que presentan les ha permitido colonizar todos los hábitats
acuáticos. Sacabambaspis es un género extinto de peces
agnatos (sin mandíbulas) que vivió en Período Ordovícico Superior.
Está relacionado con el género Astraspis. Su tamańo
era de aproximadamente 30 centímetros. Su cabeza era ancha, con los
ojos muy juntos en la parte delantera. Las partes superior e
inferior de la coraza de la cabeza estaban demarcados por unas 20
placas más pequeńas a cada lado, entre las cuales se escondían las
agallas. El cuerpo, ahusado, acababa en una única aleta caudal que
se extendía más allá de otras dos aletas, dorsal y ventral, y en una
extensión del notocordio con una pequeńa aleta al final. Al carecer
de aletas direccionales, es probable que no fuera un buen nadador.
Tenían fuertes escudos óseos en la cabeza que les protegerían de los
depredadores y estaban cubiertos de escamas de diversos tipos; en
cambio las especies de agnatos actuales tienen apariencia de
anguila, carecen de escamas y tienen una piel mucosa y resbaladiza.
Sacabambaspis janvieri vivió en aguas costeras de un
mar poco profundo que se extendía por parte de América del Sur. La
posición y la acumulación de los fósiles hallados sugiere que los
peces murieron por una súbita corriente de agua dulce que redujo la
salinidad del agua por debajo de su nivel de tolerancia. Algunos
fósiles de Sacabambaspis janvieri provienen de la
formación La Cantera, Precordillera de San Juan, Argentina. En el
país vecino de Bolivia, se han recuperado muy buenos ejemplares
atribuidos a este género de pez.
Cordylodus proavus.
Müller, 1959.
|
 |
|
 |
|
pieza dentaria
de Cordylodus proavus, Precordillera de la
Provincia de San Juan, Argentina. (*). |
|
Aspecto
de Cordylodus proavus (*) |
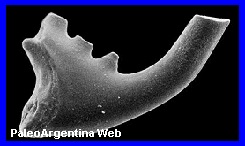
Ordovícico.
Es un género extinto de conodontes en la familia de
Cordylodontidae. Los conodontos son
una clase de cordados marinos extinta. Durante muchos ańos se
conocieron solo a partir de microfósiles dentiformes de
escasos milímetros que, a pesar de su abundancia, había sido siempre
encontrado de forma aislada. Se encuentran en el registro fósil
desde el Cámbrico medio hasta la base del Jurásico, pero
Cordylodus proavus proviene del Ordovícico exclusivamente. A
principios de la década de 1980 se hallaron por primera vez
conodontos asociados con el fósil del organismo al que
pertenecieron, en un yacimiento de Konservat-lagerstätten. Ello es
debido a que el cuerpo de estos animales era blando y solo los
dientes habían fosilizado en la mayoría de casos. En 1994 se
hallaron fósiles muy bien conservados del género Promissum,
sorprendentes por su gran tamańo y con caracteres similares a
Cordylodus proavus. A partir de impresiones de
conodontos conocidas, denominadas a veces condontóforos, se deduce
que eran organismos de aspecto similar a una anguila con 15, más
raramente 19, elementos en la cabeza dispuestos con simetría
bilateral, incluyendo un aparato alimentario radicalmente diferente
de la mandíbulas de los modernos peces. A pesar de su feroz
apariencia, se cree que fueron animales herbívoros; los "dientes"
eran usados probablemente para filtrar plancton. No obstante, no es
imposible que algunos usaran los dientes para morder. Tal vez las
tres morfologías dentarias (coniforomes, ramiformes y pectiniformes)
tenían diferentes funciones. La posición lateral de los ojos no es
común en animales carnívoros. Hoy está ampliamente admitido que los
conodontos poseían grandes ojos, aletas con radios, músculos en
forma de galón y notocordio. Estas tres últimas características son
definitorias del filo cordados (por ejemplo, vertebrados) según las
clasificaciones más recientes. Se han encontrado fósiles de
Cordylodus en Australia, Canadá, China, Colombia,
Kazajstán, Malasia, México, Noruega, Rusia, Suecia, Alaska y Estados
Unidos . En nuestro país, se han encontrado Conodontos en las
cuencas sedimentarias del Oeste y Noroeste de Argentina. De las
numerosas formas identificadas se destaca la presencia de las
especies: Cordylodus proavus, Paroistodus proteus, horridus
horridus, Yangtzeplacognathus crassus, Histiodella sinuosa,
Prioniodus elegans, Oepikodus evae , Baltoniodus navis ,
Eoplacognathus variabilis , Eoplacognathus suecicus, Pygodus serrus,
Pygodus anserinus y Amorphognathus superbus.
Particularmente Cordylodus proavus proviene de Cerro
La Chilca, Precordillera de la Provincia de San Juan, Argentina.
Macrocystella durandi.
Aceńolaza, 1999.
|

|

|
|
Fósil de
Crinoideos primitivos Macrocystella sp. Imagen de
archivo (*). |
Aspecto viviente de Macrocystella
sp. (*) |
Ordovícico. Lirio de
mar o crinoideo extinto de la clase Rhombifera (extinto). Los Lirio
de Mar, es el nombre común de un equinodermo que vive sobre
todo en los mares tropicales. Tiene el cuerpo en forma de disco
cubierto de placas óseas y brazos plumosos que se extienden hacia
arriba desde el cuerpo formando una copa. Tanto la boca como el ano
del animal se encuentran en la cara superior del disco corporal. En
la cara inferior de éste hay un tallo o pedúnculo con el que el
animal se fija al fondo marino. Algunos lirios de mar lo pierden al
alcanzar la madurez y usan los brazos para nadar o arrastrarse por
el fondo del mar. Las formas que permanecen fijas al fondo durante
toda su vida viven sobre todo en aguas profundas. Los lirios de mar
tienen colores brillantes. Suelen considerarse la
forma más primitiva de los equinodermos. En la actualidad sólo
sobreviven unos pocos centenares de especies, pero se han
encontrando miles de fósiles, en especial en las calizas originadas
durante el paleozoico. En Argentina se han encontrados estos
organismos en casi todos los periodos geológicos, pero en el
Paleozoico los restos mas significativos de Macrocystella
provienen de las localidades en la Cordillera Oriental (Andes
argentino-bolivianos). La totalidad del material proviene de los
niveles pelítico arenosos de las Formaciones Casa Colorada, Al
farcito, Rupasca y Saladillo, y afloran en
diferentes localidades de la Quebrada de Humahuaca, en la provincia
de Jujuy, y en el
Angosto de La Quesera de la provincia de Salta.Se considero una
posible migración de Macrocystella desde el margen gondwánico
sudamericano hacia la región mediterránea y se discute la taxonomía
del género.
Lingulocystis
elongata.
Thoral 1935.
|
 |
|
 |
|
Fósil de Lingulocystis.
(*) |
|
posible
aspecto de Lingulocystis. (*). |
Ordovícico. La evolución de
los equinodermos comenzó durante la época cámbrica, convirtiéndose
en elementos comunes en los mares del Ordovícico de latitudes medias
y altas. El registro de equinodermos ordovícicos en la Argentina
está limitado a los pelmatozoos. Los fósiles suelen aparecer
fragmentariamente, siendo escasos los ejemplares completos. Es bien
conocido los grandes afloramientos de rocas sedimentarias del
Ordovícico en Argentina, así como su abundante contenido
fosilífero. Hay que destacar las secuencias clásticas cambro-ordovícicas
gruesas, de más de 9.000 metros, en los afloramientos de la
Cordillera Oriental del noroeste argentino, y las secuencias calizas
cambro-ordovícicas que caracterizan la Precordillera del occidente
argentino. El cuerpo de Lingulocystis se puede dividir
en tres sectores, un tallo largo, una teca aplanada y varios
braquiolos biseriales que se elevan desde el margen superior de la
teca. El tallo alcanza hasta 20 centímetros de largo, formado por
numerosas columnas crenuladas cilíndricas de grosor desigual. Eran
filtradores, y se alimentaban de zooplancton, como foraminíferos,
pequeńos crustáceos y moluscos, y fitoplancton. Cuatro muestras de
Lingulocystis elongata bastante completas de teca con
tallo fragmentario, una de ellas con braquiolos adheridos y varios
trozos aislados de teca y tallo, provienen de las lutitas de la
Formación San Bernardo aflorando en la Sierra de San Bernardo,
Provincia de Salta.
Notopeltis
orthometopa. Harrington 1938.
|
|

|

|
|
Trilobite
Leonaspis sp |
Trilobite
Phacops |
Trilobite
Calymene |
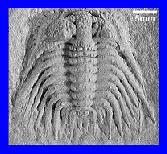
Cambrico.
Nombre común de una clase de artrópodos marinos extintos. Medían
desde unos pocos milímetros hasta casi 65 cm de largo, aunque
la mayoría de las especies tenía entre 3 y 7 centímetros.
Vivieron durante el paleozoico (iniciado hace 570 millones de ańos
y finalizado hace 245 millones de ańos), predominando en la primera
parte de esa era. Su nombre se debe a la disposición de su
exoesqueleto o caparazón externo en tres lóbulos. El exoesqueleto,
la parte del organismo mejor conservada, estaba constituido por una
sustancia dura llamada quitina, que cubría el dorso del animal.
Los trilobites tenían dos ojos compuestos que, en algunas especies,
presentaban unas lentes muy comprimidas y podrían haber servido
como dispositivo de alarma sensible a la luz para detectar el
movimiento. En otras especies los ojos tenían un menor número de
lentes, aunque más complejas, que quizá eran capaces de formar imágenes
y percibir la profundidad. Estos invertebrados vivían en vertientes
y plataformas alrededor de los márgenes continentales y en los
mares poco profundos que cubrían áreas del planeta que hoy son
masas de tierra. La mayoría habitaban en el fondo, aunque algunos
podrían haber flotado o nadado: por ejemplo, aquellos que poseían
unos ojos muy grandes con un campo de visión amplio, como
Carolinites, podrían haber sido nadadores que habitaban en aguas de
superficie. Otros, con los ojos reducidos o sin ellos, preferirían
las aguas más profundas y oscuras. Muchos de ellos, como Olenellus,
horadaban el fondo del mar para protegerse y buscar alimento. Los
trilobites emplearon una gran variedad de estrategias de alimentación.
Muchos surcaban el fango del fondo de los mares y océanos
ingiriendo los sedimentos para filtrar la materia orgánica. Otros
fueron depredadores o carrońeros. La mayoría podía adoptar una
posición defensiva enrollándose sobre sí mismos, de modo que sólo
estuviera expuesto el exoesqueleto. Los restos fósiles de
trilobites son útiles porque ayudan a los científicos a
desarrollar las escalas de tiempo relativas de los ecosistemas
marinos primitivos. Ya que estos animales evolucionaron muy rápido
y estaban ampliamente distribuidos, se pueden comparar los fósiles
encontrados en las distintas capas de roca de diferentes regiones
para estimar la edad relativa de las rocas. En este aspecto, los fósiles
de trilobites son sobre todo útiles para desarrollar las escalas
temporales de la era paleozoica inferior. En la Republica Argentina,
se pueden hallar fósiles de Trilobites del Cambrico en rocas de la
Provincia de San Juan y Mendoza (Cerro Martillo) pero también se
han descubierto fósiles bien preservado y de gran tamańo en la
Provincia de Salta. En las Islas Malvinas se han identificado
los géneros
Edelsteinaspis, Namanoia y Chondrinouyina.
Phacops orurensis.
Bonarelli, 1921.
|
 |
|
 |
|
Ejemplar de Phacops
en el Museo Paleontológico de Zapala. |
|
Aspecto de Trilobites
del genero Phacops. |
Devónico. Es un
género extinto de trilobites facópidos que vivieron durante el
Devónico. Su nombre significa "ojo de lente", debido a sus ojos
formados por pequeńos lentes de calcita. Vivía en mares cálidos y
poco profundos, siendo encontrado en todo el mundo. Su dieta se
basaba en partículas orgánicas. Phacops era un
trilobites que era depredado por varios peces, por lo que contaba
con doce placas de su tórax para proteger sus patas y la parte
dorsal. Un abultado escudo protegía su cabeza, y sus ojos tenían
lentes duros de calcita. Como mecanismo de defensa, este trilobites
se enroscaba sobre sí mismo, de modo que fuera imposible su ingesta.
Medía aproximadamente 4,5 cm. Géneros relacionados: Calymene
y Cheirurus. Especies relacionadas: P. rana,
P. cristata, P. schlotheimi, P. speculator, P. stokesi, P.
recurvus, P. latifrons y P. brocki.
Cruziana
semiplicat.
Salter, 1854.
|
 |
|
 |
|
Un buen ejemplo de icnofosiles de Cruziana. Comunes
en el Cerro San Bernardo de Salta. (*). |
|
Imagen
ilustrativa del icnofosil y el organismo que las produjo
durante su
reptación.
(*). |
Devónico -
Silúrico.
Es un icnogénero de icnofósil (huella de actividad de un ser vivo)
que corresponde a las huellas de reptación de trilobites en los
fondos marinos del Paleozoico (hasta el Devónico), desde hace de
unos 570 a 360 millones de ańos atrás.
Hoy constituye uno de los fósiles emblemáticos en la estratigrafía
del Paleozoico Inferior mundial, y común en afloramientos con
centenas de ejemplares extraordinariamente preservados en paisajes
deslumbrantes de la Cordillera Oriental Argentina. En
el Cerro San Bernardo atesora una larga evolución geológica. Hace
500 millones de ańos, lo que hoy es el espacio de Salta era una
plataforma marina, cubierta por un mar somero cercano a la costa. En
esas aguas vivían una gran variedad de invertebrados como los
trilobites, graptolites, bivalvos, braquiópodos, cistoideos,
ostrácodos, y otras formas de vida que desaparecieron para siempre
de la faz de la Tierra. Al morir, sus caparazones quedaban
sepultados en el fino fango del fondo, donde con el correr del
tiempo se fosilizaron, pero también sus trazas o huellas. La
abundancia de fósiles indica que el agua tenía buena oxigenación y
movilidad, mientras que los trilobites presentes serían evidencia de
que era un tanto fría. Las marcas de oleaje y fósiles rotos
acumulados cerca de las antiguas playas por fuertes tormentas,
pueden verse en algunos cortes del camino a la cumbre. También se
conoce en la Formación Los Espejos en el Silúrico de San Juan y en
otras regiones de Argentina.
Argentinaster
bodenbenderi.
Ruedemann, 1916.
|
 |
 |
|
Argentinaster bodenbenderi.
Devónico en la
precordillera en la Provincia de San Juan. Museo Paleontología de la
Universidad Nacional de Córdoba. |
Aspecto de
un Ophiuroidea.
(*) |
Devónico.
Los ofiuroideos (Ophiuroidea)
u ofiuras son una clase del filo Echinodermata (equinodermos). Presentan
simetría pentarradial y tienen un aspecto parecido a las estrellas de mar
(asteroideos), con cinco brazos que salen de un disco central. Seguramente,
ofiuroideos y asteroideos son grupos hermanos. Tienen el cuerpo pequeńo y
aplanado formado por un disco redondeado por cinco brazos articulados muy finos
y largos que surgen bruscamente del disco central y pueden estar ramificados.
Cada uno de los brazos consiste en una columna de osículos (llamadas vértebras),
articulados entre sí, unidos por músculos y cubiertos por placas. Tienen los
surcos ambulacrales cerrados, pero con orificios para que salgan los pies
ambulacrales, que son lisos, sin ampollas ni ventosas, pero con una musculatura
muy desarrollada. Se han encontrado ejemplares muy bien preservados en el
Devónico (entre 416 – 359 millones de ańos en la precordillera en la Provincia
de San Juan.
Ophiuroidea.
Green, 1895.
|
 |
|
 |
|
Fósil Ofiuroideo, una
variedad de estrella de mar de la Formación Fox Bay.
Devónico de Islas Malvinas. |
|
Aspecto en
vida de Ofiuroideo,
|
Devónico.
Es una
clase dentro de los equinodermos muy estrechamente relacionados con
los asteroideos o estrellas de mar comunes. Son animales
invertebrados que caracterizan por contar con 5 brazos unidos a un
disco central, por lo que mantienen su estructura secundaria
pentarradial. La diferencia más aparente respecto a los asteroideos
es que los brazos tienen una anchura menor respecto al tamańo del
disco por lo que se pueden observar espacios que separan un brazo
del siguiente. La morfología de los ofiuroideos es similar a otros
equinodermos pero con una simetría radial más marcada que ningún
otro. El disco es la parte que contiene todas las vísceras del
animal incluyendo los sistemas digestivos y reproductivos, que en
otras clases se pueden encontrar en los brazos. Ofiuroideos han sido
encontrados en la Formación Fox Bay, en las Islas Malvinas.
Plerochonetes sp.
|
Molde y contra molde del
braquiopodo Plerochonetes (=Lomoella sanjuanina) del
Devonico temprano (416-398 millones de ańos), procedente de
la precordillera de San Juan.
Museo Paleontología de la Universidad
Nacional de Córdoba. |
Devónico.
Son invertebrados marinos, bentónicos,
gregarios y filtradores, que tienen el cuerpo protegido por dos
valvas calcáreas (en algunos son quitinofosfáticas), perforadas, que
se articulan entre sí mediante una bisagra o charnela (parecida a la
de los Bivalvos, aunque mucho más simple). Las valvas
se abren mediante un complejo sistema de músculos que permiten la
entrada del agua que será filtrada por un órgano especial, llamado
lofóforo.
Los Braquiópodos se desarrollan desde el Cámbrico hasta nuestros
días. Durante el Paleozoico fueron uno de los filums más
importantes, aunque con la crisis Permico-Triásica se extinguieron la
mayoría de las especies. A pesar de ello, durante el Mesozoico y en
especial en periodo Jurásico, resurgen con fuerza a partir de este
periodo, comenzar un declive que llega a la actualidad, donde se
conocen unas 70 especies. En Argentina se han encontrado en el
Devonioco temprano, es decir, entre 416 y 398 millones de ańos antes
del presente, en la zona de la precordillera de la Provincia de San
Juan. También se conoce el braquiópodo Costuloplica sp.,
un braquiópodo de unos 300 millones de ańos, procedente de la
provincia patagónica de Chubut.
Spirifer
hawkinsii. Morris &
Scharpe, 1846.
|
|
|
Fósiles
de braquiópodos Spirifer hawkinsii recolectados por Charles
Darwin en las Islas Malvinas durante el viaje del Beagle el
22 de marzo de 1833. Museo de Historia Natural de Londres. |
Devónico.
Los braquiópodos son un filo de animales
marinos pertenecientes al clado de los lofoforados. Aunque se han
descrito más de 16 000 especies fósiles, solo existen unas 335
especies actuales. Aparecen en el registro fósil desde el Cámbrico
Inferior.Son animales con dos valvas que están unidas en la región
posterior. El sistema muscular es conformado por los músculos
aductores y diductores, que le confieren movilidad a las valvas,
permitiendo que se abran, de esta manera pueden filtrar el agua en
el lófoforo, y capturas las partículas nutritivas que son su
alimento. El aparato genital (gónadas), es simple, existiendo una
diferenciación sexual. Los braquiópodos viven sujetos al sutrato, ya
sean fijos mediante una masa muscular larga denominada pedúnculo que
se une a la valva mayor o peduncular (articulados), o que se une a
las dos valvas (inarticulados), otros en cambio posan libres en el
lecho marino. Si bien en distintas partes del territorio argentino
se han encontrado braquiópodos, esta especie en particular de
Spirifer hawkinsii fue recolectada por Charles Darwin en las
Islas Malvinas.
Undichna bina. Anderson 1976.
|
 |
|
 |
|
El rastro fósil Undichna de la
Formación Brenton Loch. Devonico de Islas Malvinas. |
|
Posible
aspecto del pez sin mandíbulas Undichna, que dejo sus
rastros en la parte onda de un lago salobre. |
Devónico. Es un rastro fósil de las aletas de pez dejadas
como impresión fósil en un sustrato mientras se desplazaba, o la
impresión opuesta en un sustrato suprayacente; este tipo de fósil es
un icnofósil. En este caso es un icnogenero específico, Undichna ; el
término "undichna" significa traza de onda. En la actualidad, los Undichna más
antiguos conocidos fueron hechos por cefaláspidos , que solo tenían
escenarios presuntivos de movimiento, debido a
la forma fisiológica de los cefaláspidos como precursores de
los teleósteos (peces óseos). Se han encontrado enormes colecciones
en la frontera de Inglaterra y Gales , en un entorno de cauce
antiguo. Las impresiones en forma de coma sugieren que los
osteostracanos usaban sus primitivas aletas pectorales en forma de
paleta para apuntalar su casco antes y durante el "despegue". El
entorno de la creación de las trazas no era marino, si salobre y la
estrecha similitud de la icnofauna de las Malvinas con la de la
sucesión Pérmica en la provincia de Eastern Cape, Sudáfrica es muy
similar.
Palaeanodonta dubi.
Amalitsky 1892.
|
 |
|
 |
|
Palaeanodonta sp. aff. P
dubia de la Formación Brenton Loch en Rory's. Islas
Malvinas, Argentina. |
|
Aspecto de una
laguna salobre en el Devonico de las Islas Malvinas,
Argentina. |
Devónico.
Una especie no marina que también se encuentra en la sucesión
pérmica de la cuenca del Karoo, Sudáfrica, y se cree que es
indicativa de una edad del Pérmico Medio. Pero Palaeanodonta
también fue hallado en las Islas Malvinas, Argentina. Como otros
bivalvos carecen de cabeza diferenciada y de tentáculos. No tienen
maxilas ni rádula. La boca presentaría palpos labiales carnosos.
Respiraban mediante uno o dos pares de branquias, generalmente
laminares. Son invertebrados, es decir que no tiene huesos. Todos
los representantes de esta clase son acuáticos, tanto marinos como
dulceacuícolas, y pueden encontrarse desde los límites superiores de
la pleamar hasta las zonas abisales. Palaeanodonta,
aparece asociado a otros organismos de agua salobre.
La protección de las conchas permite que algunas
especies especializadas soporten las condiciones de la franja
costera intermareal. Típicamente, las especies que viven sobre
sustratos blandos como fangos y arenas, presentan un pie que les
permite excavar y tiene forma de hacha.
Tornoceras
baldisi. Leanza,
1968.
|
 |
|
 |
|
Fósil del
ammonoides Tornoceras (*) |
|
Aspecto de
Tornoceras (*) |
Devónico. Es una
especie de cefalópodos ammonoides que forman el orden Goniatitida.
Vivió durante el Devónico medio hace unos 390 millones de ańos.
Ecológicamente, las goniatitas se limitaron a ambientes de salinidad
marina normal, como parece ser el caso de todos los cefalópodos a lo
largo de su historia. Las goniatitas son mucho más abundantes y
especiadas en los sedimentos que representan los mares epicontinentales que
en los que representan el océano abierto. Dentro de estos mares
interiores, la mayor abundancia y diversidad de goniatitas parece
haberse logrado en entornos costeros. Los incidentes conocidos cerca
de la costa, como las lagunas, se han atribuido generalmente
al arrastre de conchas de aguas marinas. Conchas fuertemente
involutas, con una sutura que forma de seis a diez lóbulos.
Tornoceras, en encuentran en depósitos del Devonico
superior de lugares tan dispares como América del Norte, Europa,
Asia, África y Australia. En Argentina, se los ha encontrado en la
pendiente occidental de la precordillera, en la quebrada de Chavela,
en el lado izquierdo del rió Blanco, en la provincia de San Juan. En
el mismo nivel, se registraron los braquiópodos Australocoelia
y Australospirifer.
Cooksonia caledonica.
Edwards 1970.
|
 |
|
 |
| Fósil de Cooksonia
sp (*) |
|
Aspecto en
vida de Cooksonia. (*) |
Silúrico y Devónico.
Es un género de plantas extintas pertenecientes a la clase Rhyniopsida.
Fue una de las primeras plantas terrestres. Apareció a mediados del
periodo silúrico abarcando desde los 428 millones de ańos hasta los
423 millones de ańos en yacimientos de todo el mundo. La primera Cooksonia fue
descubierta y nombrada en 1937 por el paleobotánica William Henry
Lang, de la Universidad de Mánchester, Inglaterra, quien nombró la
especie en honor a su compańera de trabajo, la australiana Isabel
Cookson. En la actualidad se conocen 7 especies asignadas al
género y aunque aún no se tienen suficientes datos para afirmarlo,
es posible que no todas estén relacionadas entre sí En los extremos
de los tallos del esporófito se localizaban
los esporangios caracterizados por ser planos o ligeramente cóncavos
en su superficie a diferencia de otras especies de su género. En su
unión al esporangio el tallo se ensancha paulatinamente no
diferenciándose el paso de uno a otro. Investigaciones recientes
realizadas a partir de unos restos excepcionalmente conservados de 5
tallos unidos mediante una estructura basal han permitido establecer
hipótesis sobre el ciclo de vida de Cooksonia paranensis.
Se han hallado fósiles de Cooksonia en la Formación
Lipeón (Silúrico Superior y transición siluro-devónica), en las
Sierras Subandinas Occidentales, provincia de Salta, en Cochabamba y
Tarija, Bolivia y en las Islas Malvinas.
Chonetes falklandicus.
Morris and Sharpe,
1846.
|
 |
|
 |
|
Chonetes sp.
Ilustrativo. (*). |
|
Abajo a la
derecha, Chonetes sp en su ambiente. (*): |
Silúrico y Devónico.
Género y especie de braquiópodos extintos, o conchas de lámparas,
que se encuentran como fósiles en rocas marinas desde el Silúrico
hasta el Pérmico (alrededor de 444 millones a 299 millones de
ańos). Los Chonetes y las formas estrechamente
relacionadas fueron el grupo más longevo de los braquiópodos
productidos. La concha es pequeńa, una mitad cóncava y la otra
moderadamente convexa. El margen horizontal del caparazón tiene
espinas cortas y en ángulo que el animal usó para anclar o
adherirse. La estructura interna de la concha de Chonetes es
distintiva. En la precordillera de San Juan se registra
Chonetes fuertensis, mientras Chonetes falklandicus,
procede de las Islas Malvinas.
Australocoelia tourtelotti.
Boucot y Gill 1956.
|
 |
 |
|
Fósil de
Australocoelia sp. (*) |
Aspecto
ambiental durante el Silurico y Devonico. |
Silúrico y Devónico.
Los más antiguos integrantes de este orden fueron descubiertos en
rocas Ordovícicas y son el segundo grupo más exitoso de
Braquiópodos, cuyos descendientes habitan aún los océanos de la
tierra. Fueron el primer grupo en articular las valvas mediante
dientes y fosetas. Por lo general las conchas son bulbosas y la
comisura tiene forma de zig zag. Son braquiópodos exclusivos del
devónico, se caracterizan por sus valvas poco curvadas y una serie
de costillas muy bien definidas. Estos fósiles casi siempre se
encuentran en bloques de rocas que contienen numerosos ejemplares,
son comunes como rodados en los lechos de ríos de la cordillera, y
en el Altiplano. Fósiles de este género fueron encontrados en
Bolivia, Perú, Paraguay, Colombia, Islas Malvinas y en la Quebrada
de Talacasto, en el Oeste de Argentina
Haplostigma furquei.
Frenguelli 1952.
 |
|
 |
|
Fósil de
Lycophyta de Haplostigma sp (*). |
|
Aspecto en
vida de las Lycophyta (*). |
Devónico. Es un género
de licopodiopsido arborescente, que se encuentra solo en la primera
parte del Devónico medio. Es una clase de
plantas vasculares herbáceas. Tienen tallos que se ramifican
dicotómicamente con hojas simples llamadas microfilas y se
reproducen por medio de esporas transportadas en esporangios en los
lados de los tallos en la base de las hojas. Aunque las especies
vivas son pequeńas, durante el Carbonífero, formas arbóreas extintas
formaron enormes bosques que dominaron el paisaje y contribuyeron
adepósitos de carbón . El análisis filogenético muestra que el grupo
se ramifica en la base de la evolución de las plantas vasculares y
tienen una larga historia evolutiva. Los fósiles abundan en todo el
mundo, especialmente en los depósitos de carbón. Los fósiles que se
pueden atribuir a Lycopodiopsida aparecen por primera vez en el
período Silúrico, junto con otras plantas vasculares. Desde el Devónico
en adelante, algunas especies crecieron grandes y parecidas a
árboles. Después del evento mundial de extinción del
Pérmico-Triásico , los miembros de este grupo fueron pioneros en la
repoblación de hábitats como plantas oportunistas. La heterogeneidad
de las comunidades de plantas terrestres aumentó notablemente
durante el Triásico Medio cuando grupos de plantas como colas de
caballo, helechos, pteridospermas, cícadas , ginkgos y coníferas
resurgieron y se diversificaron rápidamente. Fósiles de
Haplostigma sp, han sido hallados en las Islas Malvinas.
Otra especie de Lycophyta del Devónico es Haplostigma baldisii,
que proviene de la precordillera de San Juan, Argentina.
Prototaxites sp.
Dawson, 1859.
| |
|
|
 |
 |
| Fosil
de Prototaxites sp. (*). |
Aspecto del hongo
Prototaxites sp. por Skymushrooms. |
Silúrico y Devónico. Los
prototaxites son especies de hace
aproximadamente 420 millones de ańos que hace 350
millones de ańos se extinguieron, período que abarca
los periodos geológicos Silúrico y Devónico. Su
altura llegaba a alcanzar los 26 pies de altura (8
metros) y crecía adoptando una forma parecida a la
de un tronco de árbol, cilíndrica compuesta por
tubos entrelazados de tan sólo 50 micras
(millonésimas de un metro, o milésimas de milímetro)
de diámetro. Está considerado el organismo no animal
más grande de aquella época, ya que las plantas
alcanzaban tan solo los 20 centímetros de altura. El
prototaxites vivió en muchas partes del planeta. El
verdadero hongo estaba bajo tierra en forma de
filamentos y algunos teórícos, no compartida por
otros, dice que alcanzaba esta altura para poder así
distribuir sus esporas a mayor distancia. Es
considerado como uno de los organismos más extrańos
que jamás haya existido. Ya en el ańo 2001 Francis
Hueber del National Museum of Natural History en
Washington DC, publicó sus estudios exponiendo que
la falta de tejido vascular evidente lo descartaba
de que fuera una planta y la clasificó como un hongo
gigante y no fue hasta el ańo 2007 cuando Boyce en
un estudio de los isótopos de carbono C-12 y C-13
del hongo fósil comparándolo con los de fósiles de
plantas de la misma época descubrió que eran muy
diferentes entre ellos y que el prototaxites careció
de fotosíntesis donde absorbía parte o todo su
carbono de otras fuentes distintas a la atmósfera,
por lo que no era una planta (solamente absorben el
CO2 de la atmósfera) apoyando así los estudios de
Hueber de que era un hongo gigante. Se cree que
consumía suelo criptobiótico (también llamado
biocrust), suelo que contiene líquenes, musgos, y
otros hongos mucho más habitual su existencia en
aquella época que en la actual.
Clarkeia antisiensis.
d'Orbygni, 1846.
|
 |
 |
|
Clarkeia antisiensis,
precordillera de la Provincia
de San Juan. Museo de Paleontología de la
Universidad Nacional de Córdoba. |
Fósiles de
Clarkeia antisiensis. (*). |
Silúrico y Devónico.
Los
Braquiópodos son invertebrados marinos de vida
bentónica y sésil, esto es, organismos fijos que
viven adheridos al fondo del mar distintos de los
Bivalvos = almejas, debido a que su concha está
formada por dos piezas esqueletarias o valvas (una
dorsal o braquial y otra ventral o peduncular),
secretadas por el manto, las cuales componen una
estructura sólida, la conchilla, que por lo común es
equilateral (bilateralmente simétrica respecto del
plano sagital) e inequivalva (por desigualdad de
tamańo y/o forma entre las valva).
Registro de braquiópodos articulados como
Clarkeia antisiensis,
provienen del Silúrico.
El género Clarkeia
se encuentra en muchos niveles y localidades en la
Formación Los Espejos (Silúrico Medio-Tardío) del
cinturón Precordillera, centro oeste de Argentina.
La abundancia y preservación de la mayoría de las
poblaciones permite un análisis exhaustivo de sus
características externas e internas y su rango de
variaciones.
Septosyringothyris jaguelensis.
Lech, 1986.
|
 |
|
 |
|
Ejemplar de
Septosyringothyris jaguelensis
recuperado en el Carbonífero de Uspallata. Museo de Mendoza. |
|
Posible
ejemplar de
Septosyringothyris
sin procedencia especifica. Col.
Musseo Municipal Punta Hermengo de Miramar. |
Carbonífero.
Estos braquiópodos
son animales con dos valvas (una superior y una inferior, al
contrario de las valvas de los bivalvos que son bilaterales) que
están unidas en la región posterior. Generalmente son bentónicos,
viviendo fijos a sustratos duros por un pedúnculo, o enterrados en
sustratos blandos, donde excavan ayudándose de sus valvas, y forman
extensas galerías. Septosyringothyris jaguelensis, se
encuentra presente en las sedimentitas marinas del Carbonífero
Superior del Centro Oeste de Argentina, especialmente en la
Formación Agua del Jagüel, Uspallata, Mendoza. Son braquiópodos
elongados transversalmente, cuyos flancos presentan numerosas
costillas simples con espacios intercostales estrechos y
subangulosos a moderadamente anchos, además de presentar el pliegue
central y el seno lisos, y la valva ventral con el área cardinal
alta a muy alta. En la subfamilia Septosyringothyridinae se le
atribuyen los géneros Septosyringothyris e
Histosyrinx, cuyas características morfológicas distintivas
se observan en un grupo de braquiópodos fósiles ampliamente
distribuidos en las sedimentitas del Neopaleozoico de Argentina.
Archaeopteris sp.
Dawson, 1871.
|
 |
 |
Carbonifero. El término
Archaeopteris suena a dinosaurio alado, pero la
realidad es que este término hace referencia a un primitivo ancestro
de los árboles. No podemos decir que sea el primer árbol conocido,
siempre entendiendo el término árbol como un organismo vegetal
fotosintético, con tronco que se eleva varios metros del suelo. Este
honor lo sustenta
Wattieza,
pero
Archaeopteris tuvo gran relevancia a mi modo de ver en
la aparición de los primero peces de agua dulce y posteriormente en
la colonización de los tetrápodos de la tierra firme. Esta planta
arborescente se vivió desde finales del Devónico, a principios del
Carbonifero, y se extendió por todo el mundo. Otras plantas como
algas, musgos y helechos ya había colonizado tierra firme de forma
tímida, preparando el terreno para la colonización de los
artrópodos. Pero retrocedamos al origen del hábitat del
Archaeopteris.
La vida oceánica se concentraba en
el Océano Iapetus, una masa oceánica situada en el ecuador, rodeada
por 3 contienentes, Laurentia, Avalonia y Báltica. La deriva
continental hizo que estas tres masas se unieran haciendo
desaparecer dicho océano. Al disminuir el espacio vital, pasando de
un enorme océano, al contorno de un super-continente, la
competitividad por el espacio y los recurso aumentó
considerablemente. Mientras los continentes seguían colisionando,
comenzó a formarse una inmensa cadena montańosa que atravesaba el
super-continente, se trata de la Cadena Caledoniana. Se piensa que
tenía gran cantidad de picos por encima de los 8000 metros, y su
inmensidad, suponía un obstáculo para las nubes cargadas de humedad
que atravesaban el continente. Este super-continente llamado
Laurusia, era un basto desierto de areniscas, hasta que estas
montańas se formaron, ya que las nubes descargaban grandes
cantidades de agua al chocar con ella, formado los primeros grandes
ecosistemas de aguas dulces.
Lepidondendron.
Sternb., 1820.
Carbonífero.
Lepidodendron o "árbol de escamas" por la forma de su tronco,
es uno de los fósiles del período Carbonífero (hace 360 a 286
millones de ańos) es la más común de las plantas encontradas en
rocas del Pensilvaniense, de un tamańo próximo a los 40 metros, y se
suelen encontrar en fosas sedimentarias de este período. En
Peńarroya-Pueblonuevo debió existir un gran bosque de estas plantas.
Ahora solo queda el carbón. Debe su nombre a la curiosa forma de sus
escamas. Vivieron en lugares semipantanosos. Eran árboles de gran
altura. Sólo se conocen por reconstrucción. Poseían un sistema
radical muy abierto. Se ramificaban en la copa con microfilos (de 20
centímetros de longitud pero un solo nervio), en los restos de las
ramas quedan cicatrices, fundamentales para reconocer las distintas
especies de Lepidodendrales. Es lo que se conoce como cicatriz
foliar, quedando en la hoja de Lepidodendron una
cicatriz romboide. El tronco está formado por las bases de las
hojas, y sobre ellas están las hojas propiamente dichas.
Probablemente eran heterospóreos y pteridófitos. Eran gigantes
hierbas, el tronco producía muy poca madera; teniendo ayuda
estructural proveniente de una región espesa de corteza, capa rígida
que no se descascaraba como en la mayoría de los árboles modernos.
Se reproducía
por esporas.
Calamites.
Carbonífero. Es un
género fósil de equisetos que alcanzaron un tamańo similar
al de los árboles. Hace unos 300 millones de ańos, en el
Carbonífero inferior, estas plantas dominaban los
ecosistemas pantanosos de todo el mundo. Podían alcanzar 30
metros de altura y 60 centímetros de ancho. Poseían
crecimiento secundario a través del cámbium, al igual que
los árboles actuales. Hoy en día, las reservas de carbón
mundiales están en gran medida constituidas por restos de
estas plantas. Otras especies conocidas: C. cisti
y C. suckowi.
Cordaites
bifolius.
Césari S. y Hünicken, M. 2013.
|
 |
|
 |
|
Laja con fragmento de rama con hojas
de gran tamańo. Museo de Paleontología de la
Universidad de Córdoba. |
|
Aspecto de
Cordaites
bifolius
(*) |
Carbonífero.
Es un género de gimnospermas extintas, que
crecía en suelos húmedos. Eran plantas vasculares que se
reproducen por medio de semillas. Fueron similares a las
coníferas, pero con hojas grandes y acanaladas. En general,
las semillas tenían forma de corazón. Mejillones y
crustáceos de aguas salobres se encuentran a menudo entre
las raíces de estos árboles. Contrariamente a lo que ocurre
con otras variedades de plantas, las semillas de
Cordaites no son infrecuentes, ya que alcanza
grandes dimensiones. En el caso de Cordaites
bifolius, fue hallado en la localidad
de Bajo Veliz, Provincia de San Luis, en la cuenca de
Paganzo. Probablemente eran unos árboles de gran tamańo,
espatulados y aciculares, dispuestas en forma helicoidal.
Esta característica es compartida por especies del
hemisferio norte. El tronco y las ramas presentan cicatrices
vasculares alineadas horizontalmente, junto a esta planta se
hallaron órganos fértiles robustos contendiendo semillas,
probablemente de la misma especie. Estas muestras de Bajo
Veliz, proceden del Carbonífero superior – Pérmico inferior,
representa ambientes pantanosos, y alberga una rica flora,
así como insectos y diversos invertebrados acuáticos.
Megarachne servinei.
Hunicken, 1980.
|
Laja con
impronta del ex arácnido gigante del Paleozoico de San Luís.
Los últimos estudios indican que se trata de un escorpión
marino.
Paul Selden, de la Universidad de Manchester, junto al fósil
de Megarachne.
Aspecto en vida. (*). |
Carbonífero.
Megarachne servinei, que estaba registrada en el Libro
Guiness de los Récords Mundiales, era considerada como una de las
arańas más aterradoras de la era prehistórica. El experto en
arácnidos Paul Selden, de la Universidad de Manchester, asegura que
la bestia de un metro de largo era un escorpión de agua. El fósil de
la criatura fue descubierto en 1980 por el paleontólogo argentino
Mario Hunicken, quien originalmente la clasificó como una arańa que
vivió hace 300 millones de ańos.
Una de las localidades
gondwanicas mas importantes del mundo, lo conforman los
afloramientos geológicos de "Bajo Veliz", la cual se
halla emplazada en el sector noroeste de la
Sierra Grande de San Luís y ocupa una angosta depresión de más de
12 kilómetros, surcada de sur a norte por el Arroyo Cautana, en
donde si se han hallado fósiles de verdaderas arańas como
Gondwanarachne argentinensis.
También se
encuentran excelentes y abundantes restos fósiles de vegetales (las
impresiones se refieren a hojas, tallos, fructificaciones y semillas platispérmicas
con buen grado de preservación) y con frecuencia restos fósiles de
insectos y de arácnidos en muy buen estado de conservación, como el
fósil de un escorpión acuatico gigante, Megarachne
servinei que erróneamente desde su descubrimiento, había
sido catalogado como un gran arácnido. Con
la aparición de este artrópodo fósil, juntamente con la paleoentomofauna y la paleoflora resulta un hecho altamente auspicioso
desde los puntos de vista paleontológico y bioestratigráfico.
Volviendo a Megarachne, se trataba de un escorpión de
unos 70 centímetros de envergadura y con un peso probable de unos
1500 gramos. Su
alimentación consistía principalmente de pequeńos vertebrados,
como anfibios, reptiles y peces. se trataba de un artrópodo, por lo
cual, poseía
una garra larga
y dos ojos grandes, mientras que las arańas suelen tener ocho ojos
pequeńos. Además pareciera tener un cuerpo robusto o caparazón, con
arrugas o prominencias en su espalda que no se han encontrado en
ningún tipo de arańa conocida por el hombre.
Es posible que
esta criatura haya vivido en una ciénaga y usaba sus garras para
abrirse paso entre el barro. Si lo comparas con alguna especie
animal existente hoy en día, probablemente escogerías un cangrejo
largo o una langosta. Hay un
nuevo fósil de la Megarachne más grande, de 45 cm. de cuerpo y 60 cm. de extensión aproximadamente, que
está bajo la custodia del seńor Guido Pollini en la localidad de Santa
Rosa del Conlara (San Luís), emplazada a 25 km de Bajo de Veliz.
A pesar
de que no se trata de la arańa más grande que haya existido en el
mundo, es claro que es una bestia asombrosa
y
con gran valor científico.
Gondwanarachne
argentinensis. Damiani-Pinto & Hunicken,
1980.
|
 |
|
 |
|
Ejemplar de
Gondwanarachne argentinensis, procedente de
Bajo de Véliz. El Museo de
Historia Natural de la Universidad Nacional de San Luís. |
|
El aspecto de las arańas
del Carbonífero - Permico era muy similar a las actuales.
(*). |
Carbonífero
- Permico.
Durante el Paleozoico Superior, Sudamérica
estaba unida a África, Australia, India y Antártida formando
Gondwana. En este contexto en el Centro-Oeste de Argentina, no
existía la Cordillera de los Andes y el paisaje dominante eran las
Sierras pampeanas y una gran cuenca, denominada de Paganzo, cuyo
sector occidental era marino y parcialmente abiertos al océano,
mientras en el sector oriental se desarrollaban paisajes con
características continentales como las representadas en el Bajo de
Véliz. Gondwanarachne argentinensis, que
proviene de esta zona, como otros integrantes de la clase Arácnida,
el cuerpo posee dos regiones o Tagmas más o menos diferenciados, el
prosoma (o cefalotórax) y el opistosoma (o abdomen). Los apéndices
se insertan en el prosoma y son un par de quelíceros, junto a la
boca, un par de pedipalpos, a veces muy desarrollados y cuatro pares
de patas locomotoras. Carecen de antenas, y suelen tener uno o más
pares de ojos simples, en lugar de grandes ojos compuestos como los
insectos. En algunos casos esos ojos son muy eficaces para su
tamańo.
Philiasptilas hunickeni.
|
 |
|
 |
|
Ejemplar de
Philiasptilas hunickeni
de la zona de Bajo Veliz.
El Museo de
Historia Natural de la
Universidad
Nacional de San Luís. |
|
Posible aspecto de
Philiasptilas hunickeni
(*). |
Carbonífero
- Permico.
Los anisópteros son
uno de los dos infraórdenes clásicos del suborden Epiprocta. Se
conocen comúnmente como libélulas, aunque este término, suele usarse
también en un sentido más amplio para designar a todos los odonatos.
Son paleópteros, es decir, insectos que no pueden plegar las alas
sobre el abdomen. Se caracterizan por sus grandes ojos
multifacetados, sus dos pares de fuertes alas transparentes y por su
abdomen alargado. Se alimentan de mosquitos y otros pequeńos
insectos como moscas, abejas, mariposas y polillas. Su hábitat
natural se encuentra en las cercanías de lagos, charcos, ríos y
tierras pantanosas, ya que sus ninfas son acuáticas.
Philiasptilas hunickeni,
fue una
libélula que vivió durante el Carbonífero – Permico de Argentina,
aunque su pobreza en el registro fósil no ayuda a ampliar la
información disponible.
El paleoambiente en el que se depositaron los sedimentos en el Bajo
de Véliz, se corresponde inicialmente con un carácter aluvial y
cortes períodos lacustres, representados por el Miembro Cautana que
posteriormente evolucionó a un largo periodo lacustre de reducida
extensión areal, representado por el Miembro Pallero y finalmente
una larga reactivación del área, generó un nuevo ambiente fluvial,
depositando las areniscas del Miembro Lomas.
Argentinala cristinae.
Petrulevicius y Gutiérrez, 2016.
|
 |
|
 |
|
Fosi de la
libélula
Argentinala cristinae del Carbonífero inferior de la
Provincia de La Rioja. Imagen de prensa. |
|
Aspecto
ambiental del Carbonífero. (*). |
Carbonífero.
Los insectos de seis alas, a diferencia de las cuatro de las
libélulas actuales, datarían de hace 325 millones de ańos
y son los más antiguos especímenes encontrados junto a otro en
República Checa.
Sin embargo, lo más llamativo sino el nombre que
recibieron algunos de los fósiles hallados. Las
llamaron Argentinala cristinae, Tupacsala
niunamenos y Kirchnerala treintamil
por el aporte a la ciencia durante la gestión "k" (2003-2015).
Justamente
Argentinala cristinae en honor a la republica y
a la ex presidenta Cristina Fernández de
kirchner, procede del Carbonífero (318 millones de ańos) del
Cerro Guandacol,
Quebrada de las Libélulas, provincia de La Rioja. Los Odonatoptera
son un superorden (a veces tratado como un orden) de antiguos
insectos alados, colocados en los Paleoptera, que probablemente
forman un grupo parafilético. Las libélulas y los
caballitos del diablo son los únicos miembros vivos de este grupo,
que era mucho más diverso en el último Paleozoico y contenía
especies gigantescas.
Xenoptera riojaensis. Pinto
1986.
|
 |
|
 |
|
Fósil de Megasecoptero.
Imagen ilustrativa de archivo. |
|
Aspecto de Megasecoptero
Xenoptera riojaensis. |
Carbonífero.
Los megasecópteros son
un orden de insectos del Paleozoico. Estos insectos
tenían piezas bucales chupadoras, que usaban para agujerear
la cubierta de las plantas y extraer materiales de alta
calidad tales como esporas y polen. Los miembros de
Megasecoptera tenían dos pares de alas (algunos paleóteros tenían
tres), que eran aproximadamente del mismo tamańo. Las alas
se mantenían probablemente de forma horizontal, como sucede
en las libélulas (Odonata, Anisoptera). La base de las alas
tendía a ser muy delgada y peciolada, como ocurre en
los caballitos del diablo (Odonata, Zygoptera). El cuerpo
era largo y delgado. Durante su relativamente breve período
de existencia, los Megasecopteros tuvieron bastante éxito.
Se ha estimado que este orden reunió al 50% de la biomasa de
insectos en algunos lugares, aunque la evidencia disponible
podría ser desorientadora. Xenoptera riojaensis pertenece
a una nueva familia Xenopteridae, proviene de la localidad
de Malanzán, La Rioja, Argentina
Orchesteropus
atavus. Frenguelli, 1950.
|
 |
|
 |
|
Paleoicnitas
de
Orchesteropus atavus (*). |
|
Posible aspecto de
Orchesteropus atavus
(*). |
Carbonífero.
El icnofósil Orchesteropus atavus se
conoce solamente del Carbonífero de Argentina. Fue descripto
originalmente en la localidad de Huerta de Huachi, cerca de Jáchal,
provincia de San Juan y mas recientemente en Mendoza. Los
palinomorfos presentes en las capas portadoras de icnofósiles son
derivados de ambientes terrestres y no se presenta plancton marino
asociado. La icnofauna analizada es notablemente distinta a las
asociaciones de aguas salobres descriptas de ambientes marinos
marginales del Paleozoico tardío de Laurasia y Gondwana.. Las
huellas de Orchesteropus se usan como indicadores de
edad y de ambientes, pero nunca se encontró el organismo como para
conocerlo. Las marcas son de numerosas patitas con tres setas o
"dedos" y se habría desplazado realizando saltos por los fondos
fangosos, dejando rastros de las caídas con sus patas. En base a la
forma de sus patas y el andar se interpreta como similar a un
cangrejo-cacerola.
Diplichnites
gouldi. Gevers
et al., 1971.
|
 |
|
 |
| |
|
|
Carbonífero.
Es un paragénero de
icnofósiles presente en rocas sedimentarias de facies marina
profunda, lacustre, eólica o terrestre desde el periodo Cámbrico hasta
la actualidad. Los rastros de Diplichnites son
realizados por artrópodos marinos y continentales durante su
desplazamiento por sustrato arenoso. Las dos pistas
paralelas se corresponden con los dos grupos de patas
o artejos situados a cada lado del cuerpo y cada pequeńa
huella que compone esa pista a las marcas individuales del
apoyo de esas patas. Es común que en un mismo rastro
aparezca una gran variedad morfológica correspondiente a
cambios en el desplazamiento del organismo que los produce.
De este modo en organismos marinos aparecen rastros en los
que las huellas de los artejos aparecen como finos trazos
realizados durante el arrastre del artrópodo debido a
corrientes de agua. Diplichnites es un rastro
característico de trilobites, junto a otras pistas de
desplazamiento como Cruziana y trazos de
transición entre ambas.
Algunos de estos icnofósiles fueron hallados
en la Formación Agua Escondida (Carbonífero superior)
aflorante en el noroeste de La Pampa y sureste de Mendoza.
Diplichnites
cuithensis.
Briggs, Rolfe & Brannan, 1979.
|
 |
|
 |
| |
|
|
Carbonífero.
Rastro muy
común de artrópodos terrestre como la icnoespecie que se ha
asignado al miriápodo del Carbonífero Arthropleura. Las
trazas de este icnofósil están formadas por dos pistas
paralelas formadas por pequeńas y numerosas huellas
negativas o positivas, según el tipo de conservación que
presente. Las huellas pueden ser tanto circulares como
ovaladas y pueden aparecer paralelas o perpendiculares a las
trazas principales. La separación entre las pequeńas huellas
es también muy variable existiendo algunas que se espacian
entre sí varios milímetros y otras que se superponen. Eran
similares a un milpiés gigante, y vivieron en el
período Carbonífero (hace 340-280 millones de ańos). Con
una longitud de al menos entre 0,3 y 2,3 metros, son
los invertebrados terrestres de mayor tamańo conocidos de
todos los tiempos. A pesar de su apariencia aterradora,
parece ser que eran herbívoros. Habitaban bosques húmedos
donde los reptiles, anfibios e insectos abundaban, y
podrían alimentarse de los musgos y la vegetación putrefacta
que encontraban a su paso entre la maleza de helechos. Se
cree que a los depredadores podría resultarles difícil
atacarles ya que estaban cubiertos de una coraza con púas
muy cortas. Icnofosiles de este curioso milpiés, fueron
hallados en la Formación Agua Escondida del Carbonífero
Tardío de la Cuenca de San Rafael, Argentina.
Fenestella
antiqua.
d'Orbigny, 1852.
|
 |
|
 |
|
Fenestrella antiqua, un
briozoo de Sierra de Languińeo, del Carbonífero - Permico de
Chubut. |
|
Aspecto en vida de
Fenestrella. (*). |
Carbonífero.
Genero de briozoos, cuyo esqueleto de las colonias de Fenestella
consiste en ramas rígidas que están interconectadas por barras
transversales más estrechas. Entre dos y ocho individuos de la
colonia habitan en cada lado opuesto de las aberturas
aproximadamente rectangulares entre las ramas (o fenestule) en una
fila, y el vacío que dejaron cuando murieron, puede
reconocerse como dos filas de pequeńos poros con borde ( o
aberturas) en el frente de cada rama. En especímenes bien
conservados, estos están cerrados por tapas perforadas centralmente.
Son briozoos, el nombre del grupo ("animales musgo")
se debe a que muchas veces su aspecto recuerda una cubierta
subacuática de musgo. Los briozoos filtran el agua y se alimentan
de minúsculos organismos. Prefieren aguas no contaminadas, quietas y
sin corriente, como las de los lagos pequeńos. Son un filo de
pequeńos animales coloniales, que presentan un lofóforo, corona de
tentáculos ciliados que sirven para captar alimento, en los que el
ano se abre fuera de dicha corona tentacular. Forman grandes
colonias que parecen corales en miniatura aunque están más
relacionados con los braquiópodos, de miembros microscópicos .
Quedan en las orillas cuando hay vientos fuertes o actividad en el
lago. Su rango se extiende desde el ordovícico hasta el periodo
actual. Las colonias tienes formas variables algunas son
incrustaciones en forma de lamina sobre conchas o piedras, mientras
que otras se desarrollan como pequeńos árboles o frondas en forma de
red. Cada colonia cuenta con miles de individuos interconectados
(zooides) cada zooide tiene un esqueleto tubular o en forma de caja.
Briozoos y similares reticulados vivían a base de filtrar alimentos
a partir de la corriente de agua que ellos mismos producían y que
fluían en una dirección única a través de los agujeros de la
colonia.
Cyathospongia.
Okulitch, 1935.
 |
|
 |
|
Aspecto de un
fósil de Cyathospongia o Arqueociatido. Ilustrativo (*). |
|
Aspecto del
ambiente marino con la presencia de Cyathospongia (*). |
Carbonífero.
También conocidos como
arqueociátidos ,
fueron animales pequeńos, de varios centímetros, y que poseían forma
cónica o cilindro-cónica. Su posición sistemática es incierta,
habiéndose incluido con los poríferos o en un filo propio.
Recientemente se dieron a conocer nuevo materiales fósiles hallados
en la Formación Sauce Grande en Sierra de la Ventana, sureste de la
provincia de Buenos Aires. Los arqueociátidos son organismos
extintos que habitaron zonas tropicales y de ambiente marino poco
profundo, poseen un esqueleto calcáreo. El registro de
arqueociátidos en Sudamérica es muy pobre. No existen afloramientos
de calizas in situ con arqueociátidos, sólo están conservados en
bloques calcáreos dentro de conglomerados de diferentes edades,
descriptos principalmente en las Islas Malvinas y en el norte de la
Patagonia. La geología de las Sierras Australes de Buenos Aires y
sus relaciones con Sudáfrica y los Andes permite interpretar por
primera vez a estas diamictitas como un depósito de origen glacial.
Archangelskyblatta vishniakovae.
Pinto, 1972.
|
 |
|
 |
|
Fósil de una cucaracha
del carbonífero de Uruguay en forma ilustrativa. (*) |
|
Posible aspecto de
Archangelskyblatta vishniakovae (*) |
Carbonífero. Fue
una cucaracha relativamente grande del Orden Blattaria. Se
originaron y se hicieron dominantes durante el
Carbonífero. Los blatodeos incluye a las cucarachas, cutias
o baratas y a las termitas. Es un orden de insectos
hemimetábolos paurometábolos de cuerpo aplanado. Actualmente
se conocen más de 4500 especies de cucarachas y unos 500
géneros. Los primeros fósiles parecidos a blatodeos datan
del periodo Carbonífero, hace 355–295 millones de ańos. Aun
así, estos fósiles difieren de los blatodeos modernos en que
disponen de un largo oviscapto u ovipositor y son los
ancestros de los mantodeos, así como de las cucarachas
actuales. Su cuerpo es ovalado y aplanado. La cabeza pequeńa
suele estar protegida por un pronoto en forma de escudo. Sus
antenas son filiformes, sus ojos compuestos son pequeńos,
las patas largas, aplanadas y espinosas, y las piezas
bucales masticadoras. Tienen dos pares de alas, de ellos las
alas del par posterior, que son grandes y membranosas, están
cubiertas y protegidas por las alas anteriores, que son más
pequeńas y están esclerotizadas. Presentan un par
de cercos laterales en el extremo del abdomen. Las
cucarachas son generalmente omnívoras, y posiblemente
Archangelskyblatta vishniakovae también lo fuera.
Una de las características de los taxones típicos del
Carbonífero Tardío, es el patrón de nervadura general y el
contorno del ala, con cuatro venas principales y poderosas
que surgen juntas desde cerca de la base del ala. En
Argentina se lo ha encontrado en el Carbonífero superior.
Otra cucaracha del carbonífero - permico es
Anthracoblattina mendesi de Brasil o Barona
arcuata de Uruguay.
Botrychiopsis
weissiana.
Kurtz &
Frengüelli 1944.
|
 |
 |
|
Fósil
de
Botrychiopsis sp.
hallado en Bajo Veliz, en la Provincia de San
Luís. Museo Paleontológico de la Universidad
Nacional de Córdoba. |
Botrychiopsis
weissiana procedente de
la Provincia de San
Juan. Museo Paleontológico de la Universidad
Nacional de Córdoba. |
Permico.
Es
un género extinto que existía en el Carbonífero al Pérmico.
Plantas vasculares sin semilla. Eran plantas Pteridophyta
que se reproducen sólo por esporas, ya
que carecen de semillas, flores y frutos. Son las primeras
plantas vasculares y aparecen, probablemente, en el período
Silúrico superior, en la Era Paleozoica. Las pteridofitas
actuales comprenden cerca de 12000 especies que crecen
principalmente en áreas tropicales con diversidad de
ambientes, y en áreas montanas húmedas. Actualmente están
separadas en dos divisiones,
Lycopodiophyta,
que contiene a las pteridofitas más antiguas con microfilos
(hojas rudimentarias) y
Monilophyta
que reúne a las pteridófitas con
megafilos (hojas normales) y aquellas que luego de haber
adquirido
sus megafilos los
perdieron en el transcurso de su evolución. Fósiles bien
preservados de Botrychiopsis weissiana
fueron recuperados en el
Carbonífero tardío, es decir entre 318 y 299 millones de
ańos antes del presente. Algunas muestras de gran valor
paleobotanico proceden de la localidad Bajo Veliz, en la
Provincia de San Luís y también en la Provincia de San Juan.
Lycopodiopsis derbyi.
Renault
1890.
 |
|
 |
|
Tronco de licófita,
Cerro La Trampa, Chubut. Formación Nueva Lubecka.
295 millones de ańos. (*). |
|
Aspecto del Bosque
de Licofitas del Permico de Patagonia. (*). |
Permico.
Las Lycophytas o
Lycopodiophytas son las plantas, vivientes y extintas, que forman un
grupo monofilético caracterizado por poseer una alternancia de
generaciones bien manifiesta (con esporofito y gametofito de vida
libre), con el esporofito formando un cormo (vástago, raíz y un
sistema de haces vasculares que los vincula) de tipo primitivo: el
vástago consta de tallo, y en las lycopsida también de hojas de tipo
microfilo (de origen diferente de los megafilos de
Euphyllophyta, probablemente derivadas de modificaciones de
esporangios laterales, con un único haz vascular), las raíces
adventicias son originadas del vástago, las raíces tienen una
característica ramificación dicotómica. En el corte transversal del
tallo y la raíz los haces vasculares forman un patrón («estela») que
al menos en las Lycopsida, en la raíz es una actinostela y en el
tallo es derivada de la protostela con el xilema exarco, los
esporangios son de inserción lateral, y en las lycopsida crecen en
estrecha asociación con los microfilos estando ubicados en su cara
adaxial, y usualmente con forma acorazonada. Durante mucho tiempo se
las ubicó junto con Monilophyta o helechos, formando la división
Pteridophyta, que hoy se sabe que es parafilética. Hasta hace poco
tiempo se creía que las lycophytas eran tan antiguas como
Psilotum y Equisetum, pero hoy se sabe que
esos dos géneros son helechos que perdieron sus megafilos y otros
caracteres de forma secundaria (después de haberlos adquirido).
Lycopodiopsis en Argentina se encuentra desde el
Carbonífero Medio (cuenca del Paganzo) hasta el Pérmico inferior (Patagonia).
Glossopteris
damudica. Feistmantel, 1881.
Permico.
Era un arbusto o árbol del género de los pteridófitos fósiles que
caracterizó al final del Paleozoico al supercontinente Gondwana, de
tal forma es la flora existente de esta planta es el género extinto
orden de las plantas Glossopteridales. Fue un árbol característico y
dominante del supercontinente Gondwana, una gimnosperma. Se trataba
probablemente de una especie inmigrante de la franja paleoecuatorial,
descendiente de Lesieya, una planta que vivía fuera de
los ambientes pantanosos del pensilvánico. A diferencia de los
árboles de hoy en día, los hace tiempo extinguidos
Glossopteris vivían en grupos de hasta un millar por acre a
apenas 20 o 25ş del polo sur, una latitud en la que no recibían luz
solar durante la mitad del ańo. En cuanto a cómo lucían, el
Glossopteris se elevaba estrechándose hacia arriba, como un
árbol de navidad. En lugar de pinochas, tenían anchas hojas
lanceoladas que caían al suelo a fines del verano. No se sabe si
estas hojas cambiaban de color, dijo Miller, pero parece probable
que sí. Vivieron en una época en que el clima antártico era mucho
más cálido, aunque los árboles tenían también que sobrevivir a un
régimen extremo de luz solar, bastante baja durante medio ańo y
total oscuridad durante el otro medio. “Hoy en día no tenemos ningún
análogo moderno para estos bosques polares”, dijo el paleobotánico
David Cantrill, curador del Museo Sueco de Historia Natural en
Estocolmo. Los anillos fosilizados de estos árboles revelan que los
Glossopteris crecían continuamente cada verano, para
detenerse abruptamente en el invierno, como si se hubiera pulsado un
interruptor. Los modernos árboles de hoja caduca retardan su
crecimiento y luego lo detienen cuando llega el tiempo frío. Gracias
a estas agrupaciones, los investigadores pueden ahora medir la
distribución de los árboles, así como calcular sus tamańos, de modo
de aumentar la información sobre cuanta luz solar y cuanta energía
estaban disponibles, claves valiosas y poco comunes sobre el clima
pérmico. Las líneas marcadas sobre Pangea seńalan las masas de
tierra que se separarían para formar los continentes actuales.
Pecopteris
anderssoni.
Halle.
 |
 |
|
Helecho con semilla del
Permico temprano de Tasa Cuna, Córdoba.
Museo Paleontológico de la
Universidad Nacional de Córdoba. |
Pecopteris (*) |
Permico.
Es un género extinto de helecho (Filicophyta)
fósil del Pérmico. A lo largo del fronde puede haber una
fuerte variación de la forma de las pínnulas. Por un lado
están las pínnulas terminales, que frecuentemente se tratan
de un carácter taxonómico de primer orden pero que, por
desgracia, no siempre se pueden usar con fines
clasificatorios por el carácter fragmentario de los
ejemplares muestreados. Poco después del establecimiento del
género Pecopteris, se encontraron
fructificaciones asociadas con las hojas, lo que confirmó
que la mayoría de las especies eran verdaderos helechos. El
registro completo de los Pecopteris abarca
aproximadamente 40 millones de ańos.
Chelichnus duncani.
Owen, 1842.
|
 |
|
 |
|
Huellas atribuidas a
Chelichnus duncani. Tomado de researchgate.net. |
|
Posible aspecto del
tetrapodo Chelichnus duncani. Por
MarMag.2026.. |
Pérmico. Se trata de
animales muy abundantes, los cuales dejaron sus huellas de
tetrápodos. Así se llama a los animales vertebrados con
cuatro extremidades (ya sean para desplazarse o manipular).
Los anfibios, reptiles, mamíferos y aves son tetrápodos;
esto incluye a los sin patas y a las serpientes, cuyos
antepasados tenían cuatro patas. Las huellas están
preservadas en areniscas que provienen de distintas canteras
de laja ubicadas en las inmediaciones de la Sierra Pintada y
de las Sierras de las Peńas, dentro del Bloque de San
Rafael, en el sudoeste de Mendoza. La importancia
de estos registros radica en que son los más antiguos de
amniotas para Gondwana (un antiguo bloque continental que
resultó de la porción meridional de Pangea, cuando se extendió
el mar de Tetis hacia el oeste). Las
huellas identificadas más abundantes corresponden a “icnotaxones
(huellas) ya conocidos del Pérmico de Laurasia.
El análisis de los posibles productores de las huellas
muestra que los mismos representarían al menos tres taxones
diferentes de tetrápodos (que estarían) relacionados a los
mamíferos y ya estarían diversificados y ocupando ambientes
desérticos.
Asimismo, informa que las rastrilladas estudiadas de la Formación
Yacimiento Los Reyunos son unos de los pocos ejemplos conocidos de
huellas de tetrápodos preservadas en ambientes eólicos para el
Paleozoico de Gondwana, e incluso para Pangea, dado que es recién para
el Pérmico más alto que se hacen abundantes. Los autores del
hallazgo y presentación son Investigadores del
Instituto de Estudios Andinos Pablo Groeber de la UBA-Conicet, junto con
colegas de otras instituciones, han dado a conocer los interesantes resultados sobre icnitas de vertebrados que datan del
Pérmico (un período de la era
Paleozoica, hace más de 250 millones de ańos).
Chelichnus gigas. Jardine 1850.
|
|
|
|
|
 |
|
 |
|
Algunas de las huellas
de Chelichnus
gigas recuperadas en Mendoza. researchgate.net. |
|
Posible aspecto de
Chelichnus gigas (*). |
Pérmico. La importancia
de estos registros radica en que son los más antiguos de
amniotas para Gondwana.
Se caracterizan porque el embrión desarrolla cuatro
envolturas: el corion, el alantoides , el amnios y el saco
vitelino y crea un medio acuoso en el que puede respirar y
del que puede alimentarse. Ésta es una adaptación evolutiva
que, a diferencia de lo que ocurre con los anfibios,
permitió la reproducción ovípara en un medio seco y
terrestre. Otras adaptaciones son tener una piel seca y
escamosa debido a un aumento de la queratinización de la
epidermis para evitar la desecación y deshidratación; poseer
una respiración exclusivamente pulmonar y tener una
fecundación interna, con el abandono de las fases larvarias
y posterior metamorfosis. Parece ser que los primeros
amniotas surgieron durante el Carbonífero Superior a partir
de los tetrápodos reptiliformes. Transcurridos algunos
millones de ańos, dos de los linajes más importantes de los
amniotas fueron diferenciándose: por un lado nuestros
antepasados sinápsidos, y por otro los saurópsidos, de los
que evolucionaron los dinosaurios, los reptiles modernos y
las aves.
Las huellas están preservadas en areniscas que provienen de
distintas canteras de laja ubicadas en las inmediaciones de
la Sierra Pintada y de las Sierras de las Peńas, dentro del
Bloque de San Rafael, en el sudoeste de Mendoza.
(*) Imagen de autor u origen desconocido.
Ver sobre nuestras
Políticas de uso de imágenes.
|
Bibliografía Utilizada.
Aceńolaza, F.G., 1978. El
Paleozoico Inferior de Argentina según sus
trazas fósiles.
Ameghiniana
15(1-2), 15- 64. Buenos
Aires.
Aceńolaza, F.G. and Ciguel,
H., 1987. Análisis comparativo entre las
formaciones Balcarce (Argentina) y Furnas
(Brasil).
10ş Congreso Geológico Argentino,
1: 229-305. Tucumán.
Aceńolaza, F.G. and Baldis,
B.A., 1984. The Ordovician System in South
America: Correlation Chart and Explanatory
Notes.
International Union of Geological Sciences,
Publication 22, 68 pp.
Oslo.
Aceńolaza, G.F., Tortello,
M.F., Vergel, M.M. and Aráoz, L., 2001.
Icnofósiles, trilobites y palinomorfos de la
Formación Santa Rosita aflorante en la
región del Abra de Zenta (Propvincias de
Salta y Jujuy).
IV Reunión Argentina de Icnología y Segunda
Reunión de Icnología del Mecrosur.
Resúmenes, p. 22. Tucumán.
Albanesi, G.L., Esteban, S.B.
and Barnes, C.R., 1999.
Conodontes del intervalo del
límite Cámbrico- Ordovícico en la Formación
Volcancito, Sistema de Famatina, Argentina.
Temas Geológico-Mineros ITGE,
26: 521-526. Madrid
Astini, R., 2001. Mid-Late
Ordovician
Trypanites
borings in solitary
solenopora from the Argentine
Precordillera.
IV Reunión Argentina de Icnología y Segunda
Reunión de Icnología del Mecrosur.
Resúmenes, p. 26. Tucumán.
Borrello, A. 1966b Trazas y
cuerpos problemáticos de la Formación La
Tinta, sierras Septentrionales de la
provincia de Buenos Aires.
Publicación especial de la Comisión de
Investigaciones Científicas de la provincia
de Buenos Aires, Fascículo
5, 1-42, 46 pl. La Plata.
Cope, E.D. An interesting
connecting genus of Chordata (Mycterops).
Am. Nat. 1886;20:1027–1031.
D.A.T. Harper, T. Servais (2014). Early Palaeozoic
Biogeography and Palaeogeography.
Geological Society of London. pp. p.51.
Del Valle, A., 1987b. Nuevas
trazas fósiles de la Formación Balcarce,
Paleozoico Inferior de las Sierras
Septentrionales. Su significado cronológico
y ambiental.
Revista del Museo de La Plata, Nueva Serie,
Sección Paleontología,
9: 19-41. La Plata.
Esteban S.B. and
Gutiérrez-Marco, J.C., 1997. Graptolitos del
Tremadoc del Sistema de Famatina
(Argentina).
XII Jornadas de Paleontología y Vta Reunión
Internacional del Proyecto 351 PICG.
Resúmenes, 59-63. A Coruńa, Espańa.
González, P. D., Tortello, M. F.,
Damborenea, S. E., Naipauer, M., Sato, A. M.
y Varela, R. 2013. Archaeocyaths from South
America: review and a new record. Geological Journal
48(2-3): 114-125.
Harrington, H. J. and Leanza,
A.F., 1957. Ordovician Trilobites of
Argentina. Department of Geology,
University of Kansas, Special Publication
1: 276 pp.
Lawrence.
Hünicken, M.A.
A giant fossil spider (Megarachne servinei)
from Bajo de Véliz, Upper Carboniferous,
Argentina. Bol. Acad. Nac. Ciencias,
Córdoba. 1980;53:317–341.
J. F. Petrulevicius and P. R. Gutiérrez.
2016. New basal Odonatoptera (Insecta) from the lower
Carboniferous (Serpukhovian) of Argentina.
Arquivos
Entomolóxicos
16:341-358.
Krapovickas,
V., Marsicano, C.A., Mancuso, A.C., Fuente,
M.E. y Ottone, G.E. 2014. Tetrapod and
invertebrate trace fossils from aeolian
deposits of the lower Permian of central
western Argentina.
Historical
Biology.
Kjellesvig-Waering, E.N. A
taxonomic review of some late Paleozoic
Eurypterida.
J. Paleontol. 1959;33:251–256.
KU
Ichnology (ed.). «Rusophycus». Consultado el 23 de agosto de
2015.
Mángano, M.G., Buatois, L.A.
and Aceńolaza, F.G, 1996a. Icnología de
ambientes marinos afectados por volcanismo:
la Formación Suri, Ordovícico del extremo
norte de la Sierra de Narváez, Sistema de
Famatina, Argentina. Asociación
paleontológica Argentina, Publicación
Especial 4,
I
ra Reunión Argentina de Icnología.
Abstract: 69-88. Buenos
Aires.
Miller, William (2011). Trace Fossils: Concepts,
Problems, Prospects. Elsevier. pp. p.19.
Pinto, I.D; Hünicken, M.A.
Gondwanarachne, a new genus of the order
Trigonotarbida (Arachnida) from Argentina.
Bol. Acad. Nac. Cienc.,
Córdoba. 1980;53:307–315.
R.W. Frey (2012). The Study of Trace Fossils: A Synthesis
of Principles, Problems, and Procedures in Ichnology.
Springer Science & Business Media. pp. p.96.
Shear, W.A; Palmer, J.M;
Coddington, J.A; Bonamo, P.M. A Devonian
spinneret: early evidence of spiders and
silk use.
Science. 1989;246:479–481.
Tollerton, V.P. Morphology,
taxonomy, and classification of the order
Eurypterida Burmeister, 1853.
J. Paleontol. 1989;63:642–657.
Waterston, C.D. Further
observations on the Scottish Carboniferous
eurypterids.
Trans. R. Soc. Edinb. 1968;68:1–20.
Woolfe, KJ (1993). "Ambientes
deposicionales devónicos en las montańas Darwin: żmarinos o
no marinos?". Ciencia antártica . 5 (2): 211–220.
Zalba, P.E., Andreis, R.R.
and Lorenzo, F.C., 1982.
Consideraciones
estratigráficas y paleoambientales de la
secuencia basal Eopaleozoica en la Cuchilla
de las Aguilas, Barker, Argentina.
5to Congreso Latinoamericano de Geología,
2: 389-409, Buenos Aires.
Ver mas
bibliografía utilizada para hacer la presente Pagina;
AQUÍ.
Ver bibliografía de divulgación sugerida;
AQUÍ. |
|