Las
plantas son organismos que desempeñaron un papel fundamental en la
evolución de la vida terrestre. Proporcionan alimento y resguardo a
muchos otros seres vivos, producen oxígeno, regulan la humedad y
contribuyen a la estabilidad del clima.
El
estudio del registro fósil de plantas nos permite establecer cuándo
se originaron los grandes grupos de ellas (como las plantas con
flores), sus momentos de diversificación y, en algunos casos, su
momento de extinción. Además, proporciona información para
comprender la evolución en el tiempo, tanto de los ecosistemas como
del clima del planeta.
La
paleobotánica es una disciplina compartida por la botánica y la
paleontología que estudia los restos de vegetales que vivieron en el
pasado. También contempla el uso de los restos para la
reconstrucción de ambientes antiguos y la historia de la vida.
Incluye el estudio de los fósiles de las plantas terrestres, y los
autótrofos marinos como las algas.
Los
fósiles vegetales son restos de individuos que se han conservado en
las rocas sedimentarias por un proceso fisicoquímico denominado
fosilización. Por lo general sólo se conservan las partes más duras,
siendo excepcional que se conserven también las partes más blandas,
gracias a esto es posible que lleguen hasta nosotros algunas de las
partes indispensables a la hora de clasificar una planta, estas son:
las hojas, parte de sus tallos, semillas y resina fósil.
Es
infrecuente hallar los restos completos de un individuo, con todos
sus elementos, lo corriente es encontrar en el sitio una variedad de
tipos de hojas, semillas, madera en estado fósil lo que hace muy
difícil el relacionar unas con otras. Esto ha dado lugar a que cada
elemento (raíz, tallo, hoja, etc.) sea clasificado en forma
independiente como si fueran especies distintas. Las categorías
—y por extensión los
componentes de las mismas—
de esta sistemática paralela basada en elementos aislados se
denominan parataxones, para diferenciarlas de los taxones formales.
Esta disciplina nos permite deducir el clima de entonces, su
evolución y la influencia sobre otros organismos.
Tipos de fósiles vegetales.
Dependiendo de cómo se hayan preservado en el sedimento se les puede
clasificar en seis grandes grupos.
Impresiones y Compresiones:
este tipo de fosilización se da cuando los vegetales se imprimen en
el sedimento dejando su huella.
Moldes y Contramoldes:
sucede cuando las partes de las plantas con más volumen se entierran
en el sedimento. Si la materia orgánica se destruye quedará el hueco
dejado por el tronco. Si este hueco se rellena con sedimento y se
forma el contramolde.
Permineralizaciones
o mineralización: se dan cuando los tejidos quedan
impregnados de minerales como la sílice o el carbonato de calcio.
Normalmente se impregnan las células luego de la destrucción de su
contenido. También encontramos frutos y semillas mineralizadas que
son objeto de estudio dentro de la disciplina conocida como
Paleocarpología.
Carbonización: al
morir la planta en un ambiente pantanoso o reductor, en un medio con
poco oxígeno, los hidratos de carbono que constituyen la celulosa se
descomponen lentamente, desprendiendo metano y anhídrido carbónico.
Esta forma de fosilización tiene distintas etapas a través de
períodos muy largos de tiempo, formándose distintos materiales:
turba, que es el carbón vegetal menos transformado, es una sustancia
negruzca de aspecto esponjoso, lignito, hulla, antracita, que es el
carbón mineral que todos conocemos.
Ámbar y copal:
resina fósil que puede contener en su interior diversos elementos de
la flora del momento previo a la fosilización.
Polen y esporas:
son estudiados por la
Palinología. Estudia polen, esporas,
dinoflagelados y cualquier palinomórfo actual o fósil. El estudio
palinológico de polen actual contribuye a la taxonomía de plantas,
certificar calidad de mieles, predecir cosechas e investigaciones
agronómicas, estudiar la naturaleza de los contaminantes biológicos
( polen alergénico) entre otras aplicaciones. Los análisis
palinologicos son también útil a la Arqueología, Climatología,
Estratigrafía, Geología del petróleo entre otras aplicaciones.
Impresiones y Compresiones
Moldes y Contramoldes:
Permineralizaciones
o mineralización
Carbonización
Ámbar y copal
Polen y esporas
¿Que son los
Periodos Geológicos?
Son lapsos en donde los
Geólogos y los Paleontólogos han agrupado sistemáticamente la
historia de la tierra en los últimos 4 mil millones de años de
evolución geológica y biológica de nuestro planeta, donde vemos
los cambios mas significativos con explicaciones amplias y
entendibles.
La escala de tiempo geológico es el marco de referencia para
representar los eventos de la Historia de la Tierra y de la vida
ordenados cronológicamente. Establece divisiones y subdivisiones de
las rocas según su edad relativa y del tiempo absoluto transcurrido
desde la formación de la Tierra hasta la actualidad.
Los siguientes diagramas muestran la duración a escala de las
principales divisiones. El primer y segundo cronograma representan,
cada uno, subsecciones de la parte marcada con asteriscos en el que
tienen inmediatamente debajo. El tercero y último representa todo el
tiempo geológico, desde el origen de Tierra hasta la actualidad.
Era Arcaica.
Período(s): Precámbrico 4600 millones de años,
Cámbrico 542 millones de años, Ordovícico 488 millones de años. Era
Florística: Arqueofítica. Se dan los primeros indicios de la vida
que surgía en el planeta:
Monera procariota (sin núcleo celular).
Bacteria, se han encontrado registros fósiles de bacterias en rocas
terrestres con una antigüedad de aproximadamente 3.500 millones de
años.
Cyanobacteria (cyanophitas) emparentadas con
las baterías, por ejemplo los estromatolitos, importantes organismos
formadores de rocas durante este periodo.
Monera eucariota, (con núcleo celular), que
abarca organismos tanto del reino animal como del reino vegetal.
Protista, representado por los cocolithophoridos, que son algas
diminutas y esféricas, que presentan un exoesqueleto calcáreo y
también fueron importantes formadores de rocas durante este periodo.
Por ejemplo Cocolito Radiolario.
Era Paleozoica
Período(s): Silúrico 440 millones de años,
Devónico 410 millones de años, Era Floristica: Paleofítica.
En
la transición a este periodo las plantas comienzan a penetrar en
tierra firme, convirtiéndose en plantas vasculares (Cormofitas), que
poseían una raíz para absorber los nutrientes, un tejido de sostén y
brotes, ocupados en la asimilación. En el Silúrico, tienen una vida
anfibia, levantando sus brotes por encima del agua.
El
método de reproducción que utilizaba este tipo de plantas fue el de
la propagación por esporas (Pteridofitas). Representan a este grupo
las Psilophitas, que manteniendo sumergidas sus raízes, elevaban sus
brotes por sobre la superficie del agua. Estos brotes eran una
simple rama cubierta con microfolios, lo que les daba un aspecto
escamoso.
Otro representante de importancia son las Licopodiatas,
descendientes de las Psilophitas, en estas los microfolios son
reemplazados por macrofolios, los que permitían una mayor absorción
de humedad. Facilitando la independencia del medio acuático y
colonizar la tierra firme.
Carbonífero y Pérmico
Período(s): Carbonífero 360 millones de años,
Pérmico 300 millones de años. Era Florística: Mesofítica.
Una
vez que las plantas conquistaron definitivamente la tierra firme se
produjo un explosivo incremento en la vida vegetal, apareciendo una
gran diversidad de especies y en gran abundancia; es por eso que a
este periodo se le denomina Mesofítico.
Durante este periodo aparecieron especies de gran porte, grandes
árboles, pertenecientes a las Licopodiatas, que llegaron a alcanzar
alturas de hasta 40 metros y diámetros de hasta 2 metros.
Sus
principales representantes son el Lepidodendrony la
Sigilaria. Una de las adaptaciones más exitosas que
tuvieron las plantas durante este periodo fue el desarrollo de los
primeros tipos de hoja. Muy simples pero comparativamente superiores
a los macrofolios.
Aparecen durante este periodo las plantas de tallo articulado, las
que presentan un tallo hueco, con interrupciones desde las cuales en
ocasiones surgen ramificaciones secundarias. Dos especies
representativas de este grupo son: Anularia (muy similar en
aspecto a la caña) y Calamites.
También en este periodo aparecen las Licopsidas, grupo de helechos
arbóreos que alcanzaban hasta 30 ó 40 metros de altura.
Era Mesozoica
Período(s): Triásico 251 millones de años,
Jurásico 208 millones de años. Era Florística: Mesofítica.
A
comienzos del Triásico las Espermatofitas, con reproducción por
medio de semillas, inician su predominio de los ecosistemas mediante
la clase Gimnospermas, cuya reproducción es mediante semillas
desnudas.
De
estas destacan las de la subclase Coniferales, que dieron origen a
Ginkgoales –cuyo único
representante actual es él Ginko biloba–
y a las Coníferas incluyendo Araucarias, Abetos, Pinos, etc., y las
Cicadales, plantas con aspecto de palmera, con troncos globosos,
coronados en un gran penacho de hojas.
Las
Bennettitales provistas de hojas pinnadas y un tronco cilíndrico y
la peculiaridad de contar con un prototipo de flor bisexual, que más
tarde aparecerá como característica de otra clase de plantas las
Angiospermas.
Posibles herederos de los Licopodios pueden ser la clase
Coniferofitas, ya que en algunos Licopodios se producía la
heterospora que pudo prosperar hasta la formación de semillas.
Cretácico
Sistema(s): Cretácico 146 millones de años,
hasta 65 millones de años. Era Floristica: Neofítica.
Ya
antes de este periodo las condiciones son propicias para las
Angiospermas, se reproducen mediante semillas cubiertas, y a partir
de ahora logran su máximo desarrollo y diversificación. No se
produce un desplazamiento total de las Gimnospermas que perduran
hasta nuestros días.
Las
Angiospermas se dividen en: monocotiledóneas, aquellas en que su
semilla no se divide al germinar como por ejemplo las Palmáceas y en
las dicotiledóneas, su semilla si se divide al germinar como son
Abedules, Encinas, Sauces, Laureles, etc.
Terciario y Cuaternario.
Ocupa los últimos 65 millones de años. Edades Paleoceno, Eoceno,
Oligoceno, Mioceno y Plioceno (Terciario, de 55 a 2,5 millones de
años) y Pleistoceno y Holoceno (2,5 o,1 millones de años. Muchos de
los grupoide plantas actuales ya tienen representantes fósiles.
Estudio y desarrollo de la paleobotánica en la Argentina.
La
disciplina tiene un desarrollo relativamente reciente en el país,
comparado con el de otras ramas de la paleontología.
En
el siglo XIX, numerosos naturalistas europeos, como Félix de Azara
(1746-1821), Alcide d’Orbigny
(1802-1857), Charles Darwin (1809-1882) y Hermann Burmeister
(1807-1892), entre otros, comentaron hallazgos de plantas fósiles
realizados por ellos o por otros, lo mismo que hicieron hacia fin de
ese siglo exploradores locales como Francisco P Moreno (1852-1929),
Ramón Lista (1856-1898) y Jorge Fontana (1846-1920).
En
el siglo XX se destacaron la labor del berlinés radicado en Córdoba
Federico Kurtz (1854-1921), de quien la Academia Nacional de
Ciencias publicó póstumamente su Atlas de plantas fósiles de la
República Argentina; el médico y naturalista ítaloargentino Joaquín
Frenguelli (1883-1958) y el geólogo italiano Egidio Feruglio
(1897-1954), entre otros.
<<<<<Egidio Feruglio
El
despegue de la paleobotánica en la Argentina tuvo lugar en la
segunda mitad del siglo XX con la labor de Alberto Castellanos
(1896-1968), discípulo de Kurtz, y de sus numerosos alumnos, que se
cuentan entre los autores de las primeras tesis doctorales en la
especialidad escritas en el país en las décadas de 1940 y 1950,
entre ellos Pedro Stipanicic (1921-2008), Carlos A Menéndez
(1921-1976), María Bonetti (1923-2015) y Sergio Archangelsky.
Estos, junto con el botánico y palinólogo Juan Carlos Gamerro
(1923-2013), sentaron las bases para el florecimiento actual de la
disciplina en la Argentina.
A
continuación te damos un ejemplo de
fosilización de un pez junto a una hoja de árbol.
Algunas de las plantas
fósiles mas conocidas en Argentina.
A
continuación, se mostraran imágenes y clasificación por edad
geológica en forma ilustrativa sobre fósiles botánicos hallados en
Argentina. Para mayor ampliación, visite el “Periodo Geológico”
citado, para conocer la fauna contemporánea a estos.
Prototaxites sp.
Dawson, 1859.
Silúrico y Devónico. Los
prototaxites son especies de hace
aproximadamente 420 millones de años que hace 350
millones de años se extinguieron, período que abarca
los periodos geológicos Silúrico y Devónico. Su
altura llegaba a alcanzar los 26 pies de altura (8
metros) y crecía adoptando una forma parecida a la
de un tronco de árbol, cilíndrica compuesta por
tubos entrelazados de tan sólo 50 micras
(millonésimas de un metro, o milésimas de milímetro)
de diámetro. Está considerado el organismo no animal
más grande de aquella época, ya que las plantas
alcanzaban tan solo los 20 centímetros de altura. El
prototaxites vivió en muchas partes del planeta. El
verdadero hongo estaba bajo tierra en forma de
filamentos y algunos teórícos, no compartida por
otros, dice que alcanzaba esta altura para poder así
distribuir sus esporas a mayor distancia.
Archaeopteris sp.
Dawson, 1871.
Carbonifero. El término
Archaeopterissuena a dinosaurio alado, pero la
realidad es que este término hace referencia a un primitivo ancestro
de los árboles. No podemos decir que sea el primer árbol conocido,
siempre entendiendo el término árbol como un organismo vegetal
fotosintético, con tronco que se eleva varios metros del suelo. Esta planta
arborescente se vivió desde finales del Devónico, a principios del
Carbonifero, y se extendió por todo el mundo. Otras plantas como
algas, musgos y helechos ya había colonizado tierra firme de forma
tímida, preparando el terreno para la colonización de los
artrópodos. Pero retrocedamos al origen del hábitat del Archaeopteris.
La vida oceánica se concentraba en
el Océano Iapetus, una masa oceánica situada en el ecuador, rodeada
por 3 contienentes, Laurentia, Avalonia y Báltica.
Lepidondendron.Sternb., 1820.
Carbonífero.
Lepidodendron o "árbol de escamas" por la forma de su tronco,
es uno de los fósiles del período Carbonífero (hace 360 a 286
millones de años) es la más común de las plantas encontradas en
rocas del Pensilvaniense, de un tamaño próximo a los 40 metros, y se
suelen encontrar en fosas sedimentarias de este período. Es lo que se conoce como cicatriz
foliar, quedando en la hoja de Lepidodendron una
cicatriz romboide. Eran gigantes
hierbas, el tronco producía muy poca madera; teniendo ayuda
estructural proveniente de una región espesa de corteza, capa rígida
que no se descascaraba como en la mayoría de los árboles modernos.
Se reproducía
por esporas.
Calamites.
Carbonífero. Es un
género fósil de equisetos que alcanzaron un tamaño similar
al de los árboles. Hace unos 300 millones de años, en el
Carbonífero inferior, estas plantas dominaban los
ecosistemas pantanosos de todo el mundo. Podían alcanzar 30
metros de altura y 60 centímetros de ancho. Poseían
crecimiento secundario a través del cámbium, al igual que
los árboles actuales. Hoy en día, las reservas de carbón
mundiales están en gran medida constituidas por restos de
estas plantas. Otras especies conocidas: C. cisti
y C. suckowi.
Cordaites
bifolius.
Césari S. y Hünicken, M. 2013.
Laja con fragmento de rama con hojas
de gran tamaño. Museo de Paleontología de la
Universidad de Córdoba.
Aspecto de
Cordaites
bifolius
(*)
Carbonífero. Es un género de gimnospermas extintas, que
crecía en suelos húmedos. Eran plantas vasculares que se
reproducen por medio de semillas. Fueron similares a las
coníferas, pero con hojas grandes y acanaladas. En general,
las semillas tenían forma de corazón. Mejillones y
crustáceos de aguas salobres se encuentran a menudo entre
las raíces de estos árboles. Contrariamente a lo que ocurre
con otras variedades de plantas, las semillas de
Cordaites no son infrecuentes, ya que alcanza
grandes dimensiones. En el caso de Cordaites bifolius, fue hallado en la localidad
de Bajo Veliz, Provincia de San Luis, en la cuenca de
Paganzo. Probablemente eran unos árboles de gran tamaño,
espatulados y aciculares, dispuestas en forma helicoidal.
Esta característica es compartida por especies del
hemisferio norte.
Botrychiopsis
weissiana.Kurtz &
Frengüelli 1944.
Fósil
de Botrychiopsis sp.
hallado enBajo Veliz, en la Provincia de San
Luís. Museo Paleontológico de la Universidad
Nacional de Córdoba.
Botrychiopsis
weissiana procedente de
la Provincia de San
Juan. Museo Paleontológico de la Universidad
Nacional de Córdoba.
Permico.
Plantas vasculares sin semilla. Eran plantas Pteridophyta
que se reproducen sólo por esporas, ya
que carecen de semillas, flores y frutos. Son las primeras
plantas vasculares y aparecen, probablemente, en el período
Silúrico superior, en la Era Paleozoica. Las pteridofitas
actuales comprenden cerca de 12000 especies que crecen
principalmente en áreas tropicales con diversidad de
ambientes, y en áreas montanas húmedas. Actualmente están
separadas en dos divisiones, Lycopodiophyta,
que contiene a las pteridofitas más antiguas con microfilos
(hojas rudimentarias) y Monilophyta que reúne a las pteridófitas con
megafilos (hojas normales) y aquellas que luego de haber
adquirido sus megafilos los
perdieron en el transcurso de su evolución. Fósiles bien
preservados deBotrychiopsis weissiana
fueron recuperados en el
Carbonífero tardío, es decir entre 318 y 299 millones de
años antes del presente. Algunas muestras de gran valor
paleobotanico proceden de la localidad Bajo Veliz, en la
Provincia de San Luís y también en la Provincia de San Juan.
Lycopodiopsis derbyi.
Renault
1890.
Tronco de licófita,
Cerro La Trampa, Chubut. Formación Nueva Lubecka.
295 millones de años. (*).
Aspecto del Bosque
de Licofitas del Permico de Patagonia. (*).
Permico. Las Lycophytas o
Lycopodiophytas son las plantas, vivientes y extintas, que forman un
grupo monofilético caracterizado por poseer una alternancia de
generaciones bien manifiesta (con esporofito y gametofito de vida
libre), con el esporofito formando un cormo (vástago, raíz y un
sistema de haces vasculares que los vincula) de tipo primitivo: el
vástago consta de tallo, y en las lycopsida también de hojas de tipo
microfilo (de origen diferente de los megafilos de
Euphyllophyta, probablemente derivadas de modificaciones de
esporangios laterales, con un único haz vascular), las raíces
adventicias son originadas del vástago, las raíces tienen una
característica ramificación dicotómica. Hasta hace poco
tiempo se creía que las lycophytas eran tan antiguas como
Psilotumy Equisetum, pero hoy se sabe que
esos dos géneros son helechos que perdieron sus megafilos y otros
caracteres de forma secundaria (después de haberlos adquirido). Lycopodiopsis en Argentina se encuentra desde el
Carbonífero Medio (cuenca del Paganzo) hasta el Pérmico inferior (Patagonia).
Glossopteris
damudica. Feistmantel, 1881.
Permico.
Fue un árbol característico y
dominante del supercontinente Gondwana, una gimnosperma. Se trataba
probablemente de una especie inmigrante de la franja paleoecuatorial,
descendiente de Lesieya, una planta que vivía fuera de
los ambientes pantanosos del pensilvánico. A diferencia de los
árboles de hoy en día, los hace tiempo extinguidos
Glossopteris vivían en grupos de hasta un millar por acre a
apenas 20 o 25º del polo sur, una latitud en la que no recibían luz
solar durante la mitad del año. En cuanto a cómo lucían, el
Glossopteris se elevaba estrechándose hacia arriba, como un
árbol de navidad. En lugar de pinochas, tenían anchas hojas
lanceoladas que caían al suelo a fines del verano. No se sabe si
estas hojas cambiaban de color, dijo Miller, pero parece probable
que sí.
Pecopteris
anderssoni.Halle.
Helecho con semilla del
Permico temprano de Tasa Cuna, Córdoba.
Museo Paleontológico de la
Universidad Nacional de Córdoba.
Pecopteris (*)
Permico.Es un género extinto de helecho (Filicophyta)
fósil del Pérmico. A lo largo del fronde puede haber una
fuerte variación de la forma de las pínnulas. Por un lado
están las pínnulas terminales, que frecuentemente se tratan
de un carácter taxonómico de primer orden pero que, por
desgracia, no siempre se pueden usar con fines
clasificatorios por el carácter fragmentario de los
ejemplares muestreados. Poco después del establecimiento del
género Pecopteris, se encontraron
fructificaciones asociadas con las hojas, lo que confirmó
que la mayoría de las especies eran verdaderos helechos. El
registro completo de los Pecopteris abarca
aproximadamente 40 millones de años.
Dicroidium. Gothan, 1912.
Dicroidium
(Carystopermaceae) un árbol Triasico de Argentina.
Otra especie de
flora de Dicroidium de Los Rastros.
Característica de la flora del
Triasico de Ischigualasto.
Triasico.Las comunidades vegetales del Triasico de
Patagonia cambian drásticamente con respecto a aquellas del
Paleozoico superior. Esta nueva asociación florística domina esta
región al igual que en Ischigualasto. La planta mas importante es
una con forma de helecho que lleva semillas y se denomina Dicroidium.
La misma pertenece a un grupo de angiospermas sumamente primitivas
que combina una serie de características como la presencia de óvulos
con hojas sumamente divididas. Hay importantes asociaciones con
otras especies de helechos primitivos y coniferas, en fin, toda una
gama rica y exuberante la cual muy probablemente constituyeron el
principal alimento de los grandes vertebrados de este periodo.
Sphenobaiera argentina.
Frenguelli, 1946.
Plantas fósiles del genero Sphenobaiera.
Museo de La Plata.
Frondes vegetales del
Triasico superior de Cacheuta. Museo de
Ciencias Naturales y
Antropológicas Juan Cornelio Moyano de Mendoza.
Triasico.Es un
género extinto de plantas que vivieron durante el
Triásico Superior al
Cretácico. El géneroSphenobaiera se utiliza para plantas
con hojas en forma de cuña y se puede diferenciar de
Ginkgo,
Ginkgoites y
Baiera la falta de un
peciolo.
Se extinguieron hace unos 72.600 años. La
familia que pertenece a este
género no es concluyente y afinidad con
Karkeniaceae había razones
morfológicas sugeridas.
Rhexoxylon brunoi.
Artabe, Brea et Zamuner 1999.
Los troncos fósiles de la formación los
colorados, en Villavicencio- Uspallata en el Museo de
Ciencias Naturales y
Antropológicas Juan Cornelio Moyano de Mendoza.
Restos de troncos en sedimentitas triásicas del área de
Paramillos de Uspallata (Cuenca Cuyana, Mendoza). Imagen del
Inst. de Estudios Andinos.
Triasico.Es un género de
coníferas de la familia Araucariaceae. Los troncos fósiles de este
genero procede de la formación los colorados, en Villavicencio-
Uspallata, en Mendoza. Son principalmente árboles grandes con un
vástago erguido masivo, alcanzando una altura de 30 a 80 m. Las
ramas suelen ser horizontales, y bien separadas; estando cubiertas
con hojas coriáceas o aciculares. Charles Robert Darwin (1809/1882)
recorrió la provincia de Mendoza (Argentina) durante 1835, como
parte de aquella histórica travesía alrededor del mundo (1831/1836)
a bordo del Beagle, expedición dirigida por Fitz Roy. Entre sus
descubrimientos más notables, y a la vez poco recordados, se
encuentra un bosque de árboles petrificados de más de sesenta
millones de años de antigüedad.
Dictyophillum tenuifolium.
Stipanicic y Menéndez, 1949.
Dictyophillum tenuifolium, del Triásico Superior del
Paso Flores (Provincia de Río Negro). Paseo Paleontológico
de Bariloche.
Aspecto de un representante de la Familia Dipteridaceae.
Triasico.Los
helechos de la Familia Dipteridaceae constituyen uno de los grupos
sistemáticos más reconocibles en las floras mesozoicas debido a su
característica venación anastomosada. Aunque este carácter es
compartido por otros helechos (e.g., Matoniaceae y géneros
como Weichselia del Mesozoico y Polypodiaceae del
Cenozoico), de todas maneras son los más típicos y ampliamente
difundidos en el Mesozoico del mundo.
Dictyophillum tenuifolium, es una especie,
originalmente descrita por Stipanicic y Menéndez (1949) del Triásico
Medio de la Formación Barreal de la Provincia de San Juan
(Argentina). El registro fósil comienza en el Triásico tardío.
Actualmente distribuidas en India, el sudeste de Asia, el este y
sudeste de China, centro y sur de Japón, y desde Malasia hasta
Melanesia y el oeste de Polinesia (Samoa)
Frondes vegetales de
Corystospermales del Triasico de San Juan. Exhibido en el
Museo de Cs Naturales y Antropológicas de Mendoza.
Aspecto de un
bosque triasico de
Corystospermales (*):
Triasico.Este
genero y especie con abundante presencia en Cacheuta (Luján de Cuyo) en la provincia de
Mendoza, como así también en San Juan, corresponde a las conocidas
plantas Corystospermales, un orden de plantas que aparecieron en el
Pérmico y se extinguieron en el Cretácico. Se trata de un grupo
relativamente pequeño de plantas con distribución principalmente en
Gondwana. Fueron las plantas dominantes del hemisferio sur durante
el Triásico, y se utilizan en la bioestratigrafía y paleoecología de
ese periodo. Se caracterizan por tener un raquis principal bifurcado
en sus frondes. Los tallos presentaban verdadero crecimiento
secundario en "cuñas" de xilema centrífugo y centrípeto. Los órganos
de reproducción femeninos son tipo cúpula, que surgen de
braquiblastos (pequeñas ramas abortadas).
Nododendron
suberosum. Artabe y Zamuner
1991.
Ejemplares de
Nododendron suberosum. Proceden de la formación
Potrerillos, Triasico de la Provincia de Mendoza. Exhibidos
en el Museo de La Plata
Triasico.Este
taxón fue descripto e ilustrado para la Formación Potrerillos
expuesta en el cerro Cacheuta. En la Formación Paramillo, localidad
de Agua de la Zorra provincia de Mendoza, se hallaron restos afines
a Nododendron
suberosumen asociación estrecha con
Neocalamites carrereiy estructuras reproductivas (Neocalamostachys
arrondoi). La familia equisetáceas es monotípica (con un
solo género), siendo su género viviente, Equisetum
(conocido como cola de caballo), que es el único sobreviviente hasta
la actualidad de todas las equisetópsidas, las cuales tienen un
extenso registro fósil entre el Devónico y el Triasico. Los
equisetos son plantas vasculares con ciclo de vida
haplodiplonte donde la alternancia de generaciones es bien
manifiesta, con esporofito y gametofito multicelulares e
independientes, con esporas como unidad de dispersión y de
resistencia.
Baiera furcate.
Lindley and Hutton, 1837.
Improntas de
Baiera furcate. (*):
Baiera
furcate, procedente de Los Potrerillo, Mendoza y
exhibida en el Museo de La Plata.
Paleobotanica. Es
un género de gimnospermas prehistóricas en el orden Ginkgoales. Está
relacionado con los géneros Ginkgo y Ginkgoites. Se lobula en cuatro
segmentos y no tiene tallo. Los fósiles de Baiera han
sido conocidos desde el Pérmico hasta el Cretácico. Las Ginkgoaceae —una familia de Ginkgoales—
son gimnospermas arbóreas con un único representante vivo actual,
del género Ginkgo y especie Ginkgobiloba. A diferencia de las cícadas, y como casi todas
las demás gimnospermas, son árboles leñosos altamente ramificados.
Puede ser reconocido por el hecho de que tiene ramas cortas además
de ramas largas. De hasta 30 metros de altura, con una corona más o
menos asimétrica y corteza gris y surcada. Sin canales de resina.
Hojas simples, de disposición espiralada, y ampliamente espaciadas
en largas ramas cerca de las puntas de las ramas, también
ampliamente empaquetadas en ramas cortas en los árboles más viejos,
con forma de abanico, bilobadas o enteras, deciduas y amarillas
brillantes en el otoño, de venación dicotómica. En Argentina se han
recuperado fósiles en la Formación Paso Flores, en la localidad
Cañadón de Pancho, área occidental del río Collón Curá, Provincia
del Neuquén, Cacheuta y Los Poterillos en la pre-cordillera en la
Provincia de Mendoza Otras especies conocidas son; Baiera cuyana
y Baieragracilis.
Anatomía y
morfología de los tallos de Equisetum thermale. 10, 14, 15
secciones longitudinales. (*).
Impronta del
tallo de Equisetum dimorphum del jurasico de Chubut
Reconstrucción
del aspecto en vida de Equisetum thermale. (*).
Jurasico. En la
zona central del Macizo del Deseado, Provincia de Santa Cruz, en
Argentina, un equipo internacional de investigadores encontró un
fósil que pertenece a una especie vegetal que no se conocía y que
vivió en condiciones extremas hace 150 millones de años en regiones
geotermales. Los resultados del trabajo fueron publicados en la
edición de abril de la revista científica American Journal of Botany.
“El nombre de la nueva especie fósil, Equisetum thermale,
refleja el lugar donde ha vivido esta planta tan particular. En el
Jurásico, en lo que hoy conocemos como la provincia de Santa Cruz,
existía un ambiente volcánico muy activo.
Austrohamia minuta.
Escapa et al.2008.
Jurasico. Vivieron hace unos 170
millones de años. Las plantas fósiles estuvieron archivadas desde su
hallazgo, alrededor del año 2000, hasta que comencé el análisis para
mi doctorado. Y ahí se descubrió que era un nuevo género de la
familia Cupresaceae”, explica Escapa en referencia a las hojas y
conos (piñas) fosilizados a los que bautizaron Austrohamia.
Muchas características nos aseguraban que se trataba de una
conífera, como son las araucarias o pinos, y particularmente una
cupresácea, como los cipreses, pero otras tantas nos mostraban que
el hallazgo no coincidía con ningún género conocido, por eso
definimos uno nuevo”, relata Escapa. El nombre completo de la
especie es Austrohamia minuta, debido al minúsculo
tamaño de los conos encontrados, que miden apenas 2 milímetros y de
los que aparecieron ejemplares masculinos, que contienen polen, y
femeninos, con semillas.
Araucaria
mirabilis.Windhausen, 1924.
Bosque
petrificado en la Provincia de Santa Cruz, Argentina. (*).
Piñas
fosilizadas de Araucaria Mirabilis sp. Museo de Mendoza.
Aspecto de
un fragmento de piña, donde se observan las semillas. (*).
Jurasico. Esuna
especie extinta deárboles de coníferasde la Patagonia,
Argentina. Pertenecea la secciónBunya(la única especie viva
de los cuales esAraucariabidwilliide
Australia)del géneroAraucaria. Se
conocena partir de grandescantidades demadera yconossilicificadamuy bien conservadodesdeelCerroCuadradoBosque
Petrificado, incluidos los troncos de
árbolesque llegaron a100metros de
altura en la vida.El
sitio fuesepultado poruna erupción volcánica
durante elJurásico Medio, hace
aproximadamente 160
millones de años. Los fósiles de
Araucariamirabilisse encuentranen gran
abundanciaen el Bosque PetrificadoCerroCuadradode la Patagonia,
Argentina. Los
bosquespetrificados deA.
mirabilisfueron descubiertospor primera vez en1919
porel botánico alemán-argentino
AnselmoWindhausen.
Durante el jurasico gran parte de Patagonia fue afectada por la
actividad volcánica, con resultados dramáticos para los vegetales
de la región, sobre todo los árboles. El Bosque Petrificado se
encuentra en la localidad fosilífera del Cerro Madre e Hija y Cerro
Cuadrado en la
Provincia de Santa Cruz, Argentina.
Otozamites ameghinoi. Kurtz, 1902.
Jurasico.
El hallazgo de
restos de polen de esta y otras especies fueron realizadas en la
Cuenca Neuquina, en el margen del arroyo Lapa, en sedimentos que
indicarían que la región estuvo bajo agua, y cuyos restos
microscópicos quedaron preservados en las orillas de las playas
existentes en aquel periodo, las cuales pertenecían a vegetación
costera del territorio.En nuestra Patagonia
formaron comunidades muy numerosas que convivieron con coníferas
y helechos en zonas llanas y abiertas (sabanas). Sus hojas eran
grandes, con láminas enteras o divididas en foliolos alargados,
dispuestas en el extremo superior del tronco formando una
corona.
Fósiles
botánicos de la Formación la Matilde (jurasico) de la
provincia de Santa Cruz, Argentina.
Jurasico. Están
representados en la actualidad por una sola especie, Ginkgo
biloba, ampliamente cultivada en todo el mundo. Son y fueron
vegetales arbóreos con hojas características por su forma como
abanico y con las venas
divididas en horqueta (dicotómicas). Son dioicas, es decir
órganos sexuales están en individuos separados, los poliníferos
en los masculinos y los ovulíferos en los femeninos.
Durante
el periodo Jurasico gran parte de la Patagonia Argentina se
diversificaron los gimnospermas. Un grupo de ellos y que en la
actualidad tienen un represéntate considerado "fósil
viviente" posee hojas iguales a sus parientes primitivos. Es
asombroso que este tipo de árbol pudo sobrevivir durante 200
millones de años, a pesar de los dramáticos cambios geográficos y
ambientales que sufrió Sudamérica.
Paraucaria patagonica.
Wieland, 2010.
Troncos
gigantes de Araucaria mirabilis y Paraucaria patagonica del
Jurasico del Bosque de Jaramillo. (*).
Jurasico. El Bosque Petrificado de
Jaramillo, en el norte de Santa Cruz, y a 256 kilómetros de Puerto
Deseado, es uno de ellos. El lugar es, literalmente, un bosque. Sus
troncos, ya de piedra, están derribados, pero sus raíces están en el
mismo lugar en donde transcurrieron sus vidas. El sitio es
impactante, y está considerado uno de los yacimientos fósiles más
importantes del país. Se encuentran los
árboles petrificados más grandes del mundo: algunos ejemplares miden
35 metros de longitud (sin contar partes enterradas, ramas menores y
copa) y 3 metros como diámetro mayor. Hasta el momento de
petrificarse contaban con 1.000 años de edad.
Fósiles de
Cladophlebis sp, del Jurasico de Patagonia. Exhibido en
el Museo Argentino de Ciencias Naturales.
Fósil de
Cladophlebis (*).
Aspecto de
Cladophlebis (*).
Jurasico. Dentro del
reino vegetal
también hay muchas plantas que, por diversos factores a lo largo de
la historia, se han extinto. El género Cladophlebis,
el cual incluyó a varias especies de helechos de hace millones de
años atrás. El género Cladophlebis es un género
extinto de pteridofitas (pteridophytas),
nombre sofisticado para los helechos y afines. Estas plantas
antiguas habitaron la Tierra durante el período
Paleozoico tardíoy el
Mesozoico, concretamente entre 284.4
a 70.6 millones de años atrás según indica el registro fósil.
Las Cladophlebis se extendieron por ambos hemisferios,
y contaba con numerosas especies. Algunos fósiles de esta fronde
fueron recuperados en la Formación Carrizal (Triásico), depocentro
de Marayes, San Juan, cerro La Brea (Jurásico Temprano), provincia
de Mendoza.
Ginkgoites
tigrensis.
Archangelsky 1965.
Fósil de Ginkgoites
tigrensis exhibido en el Museo de La Plata.
Fósil de Ginkgoites
(*).
Cretácico. Es
un género extinto de plantas pertenecientes a la familia Ginkgoaceae. Se
han encontrado fósiles en todo el mundo, en sedimentos del Triásico,
Jurásico y Cretácico. son árboles leñosos altamente ramificados. Puede
ser reconocido por el hecho de que tiene ramas cortas además de ramas
largas. De hasta 30 m de altura, con una corona más o menos asimétrica y
corteza gris y surcada. Sin canales de resina. Hojas simples, de
disposición espiralada, y ampliamente espaciadas en largas ramas cerca
de las puntas de las ramas, también ampliamente empaquetadas en ramas
cortas en los árboles más viejos, con forma de abanico, bilobadas o
enteras, deciduas y amarillas brillantes en el otoño, de venación
dicotómica.
Pterodofitas.
Cretácico. Este grupo incluye a los vegetales con estructura vascular más
antiguos que vivieron sobre el planeta. Los Helechos conforman la
parte esencial del grupo y los más antiguos se registran en
estratos paleozoicos en todo el mundo. Durante el Cretácico en la Patagonia, los helechos
proliferaron tanto en comunidades boscosas como en las praderas y
valles. En general han pertenecido a familias que aún hoy perduran,
aunque en menor escala, como las Osmundaceas, Gleicheniaceas o
Dipteridaceas. Pero durante este período aparecieron o se
diversificaron otros helechos que hoy predominan en nuestras floras,
como las Pteridaceas, Cyatheaceas, Hymenophyllaceas, o los
"helechos serrucho" que hoy en día adornan nuestros
jardines.
Bennettitales.
Cretácico. Las Bennettitales pertenecieron al grupo de las Gimnospermas
(plantas con semillas desnudas), fueron netamente mesozoicas
(vivieron en la Era Secundaria) y se extinguieron a fines del período
Cretácico. En nuestra Patagonia formaron comunidades muy numerosas
que convivieron con coníferas y helechos en zonas llanas y abiertas
(sabanas). Se las ha considerado antecesoras de las Angiospermas
(plantas con flores) por poseer sus estructuras reproductivas
características semejantes a las flores actuales. Fueron plantas
semejantes a las Cycadales, tenían el aspecto de pequeñas palmeras
de 1 a 3 metros de altura, con troncos cilíndricos, columnares o a
veces globosos y con pocas o ninguna ramificación.
Angiospermas.
El fósil de la
angiosperma más antigua conocida. Lajas fosilíferas de la Sierra El Gigante, en
el noroeste de la provincia de San Luís.
Aspecto de las
antiguas
angiospermas.
Cretácico.
Uno de los rasgos mas interesantes de los fósiles
de Baquero, es que aparecen por primera vez en todo Gondwana las
Angiospermas, las cuales son las primeras plantas que utilizan sus
flores para reproducirse, lo que implica, la interacción con
insectos polinizadores. Antes de que aparecieran este nuevo grupo,
solo existían las coniferas de colores verdes, dando al paisaje
monocromia, pero las Angiospermas se extendieron en todos los
ambientes y formas como árboles, arbustos, plantas y pastos, los
que especialmente en la primavera Cretácica coloreaban toda la región.
Esto fue la consecuencia evolutiva que un gran grupo de animales
pudieran percibir los colores de plantas y frutos.
Araucaria grandifolia.Del Fueyo y Archangelsky 2002.
Hoja de
Araucaria lefipanensis del Cretácico
de Chubut. Museo Egidio Feruglio.
Detalles de piña o cono de
Araucaria sp (*). Araucaria fosilizada junto a la rama. (*)
Cretácico. Las
Coniferas fueron y son plantas generalmente arbóreas que alcanzan
grandes alturas y suelen vivir en comunidades puras o bien asociadas
a otros grupos de Angiospermas. Es uno de los grupos de Gimnospermas
más antiguos, reconocido ya en estratos del Carbonífero, y que
adquirió gran difusión en el Mesozoico, especialmente durante los
períodos Jurásico y Cretácico. Durante el Jurásico y gran parte
del Cretácico, fueron los árboles por excelencia como lo
demuestran los numerosos testigos petrificados que se encuentran en
diferentes latitudes del planeta. En la Patagonia, se halla uno de
los ejemplos más espectaculares conocidos en todo el mundo, el de
los Bosques Petrificados de la provincia de Santa Cruz, reconocido
como Monumento Natural.
Ginkgoales.
Cretácico. Están representados en la actualidad por una sola especie, Ginkgo biloba, ampliamente cultivada en todo el mundo. Son y fueron
vegetales arbóreos con hojas características por su forma como
abanico y con las venas divididas en horqueta (dicotómicas). Son
dioicas, es decir órganos sexuales están en individuos separados,
los poliníferos en los masculinos y los ovulíferos en los
femeninos. Los óvulos son, en la especie actual, simples
pedunculados. Su historia geológica se remonta a la era paleozoica
con el hallazgo de diferentes tipos foliares similares a los
actuales o mesozoicos, también referidos a la clase. En el Cretácico
de la Patagonia, se han encontrado importantes restos de este grupo,
con especimenes fértiles, portadores de numerosos óvulos
dispuestos en forma compacta (tipo estróbilo) y que se describieron
como género Karkenia.
Symphyomyrtus s.p.
Schauer.
Fósil de
Symphyomyrtus
procedente de la Laguna del Hunco, en la
provincia de Chubut. Imagen de prensa.
Aspecto de
Symphyomyrtus (*)
Paleoceno.Se ha logrado
determinar que unos fósiles de hojas, flores, frutos y brotes,
descubiertos en la Patagonia argentina, son de eucalipto, y datan de
hace 51,9 millones de años. La identificación de estos fósiles los
convierte en los macrofósiles de eucalipto más antiguos validados
científicamente, y los únicos reconocidos de manera concluyente como
nativos de un territorio de fuera de Australasia. Un equipo de
investigadores identificó los fósiles como pertenecientes a un
subgénero de eucalipto, el Symphyomyrtus, y esto
también demuestra que dicho subgénero es más antiguo de lo que se
pensaba hasta ahora. Los investigadores identificaron un conjunto de
estructuras características del género Eucalyptus en
la morfología (forma) de las huellas encontradas en distintos
fósiles.
Fósil de
Rhamnaceae, hallado
en la Formación Salamanca, Chubut.
Aspecto de un
Rhamnaceae actual (*)
Paleoceno.Un
equipo científico de Estados Unidos y Argentina ha identificado que
se corresponde con el grupo de las Rhamnaceae y ha podido
profundizar en cómo fueron las conexiones de la vegetación entre los
continentes. Investigadores de la Cornell University (Estados
Unidos), de la Universidad Nacional del Comahue y del Instituto de
Investigaciones en Biodiversidad y Ambiente (INIBIOMA-CONICET) de
San Carlos de Bariloche, Rio Negro (Argentina) y de la Universidad
Estatal de Pennsylvania (Estados Unidos) han estudiado flores
fósiles halladas en rocas de 64 millones de años en la denominada
Formación Salamanca, en la Patagonia (Chubut, Argentina).
Menucoa cazaui.Petriella
1969.
Tronco
fósil de Menucoa cazaui, Terciario de Rio Negro.
Exhibido en el Museo de La Plata.
Aspecto de un
bosque
de Menucoa cazaui,
(*)
Paleoceno.Su
registro fósil se extiende desde el Pérmico, hace al menos 250
millones de años a la actualidad. Las cicadáceas (nombre científico Cycadaceae) son
una familia de gimnospermas del
orden Cycadales. En apariencia son similares a las palmeras
(familia Arecaceae), pero en realidad son filogenéticamente
distantes de éstas, formando parte de la división de las cicadófitas.
Ejemplares fósiles de Cycasdatan del Cenozoico en Laurasia, aunque otros
más antiguos y posiblemente relacionados se remontan al Mesozoico. Cladísticamente están
bastante separadas de su familia hermana Zamiaceae, también
perteneciente al orden Cycadales.
Menucoa cazaui
es conocido a partir de un tronco petrificado de Cycadales, del
terciario temprano de la provincia de Rio Negro, Argentina
Raiguenrayun cura.Barreda,
V. et al.
Fósil de Raiguenrayun cura,
en el Museo Paleontológico de la Ciudad de Bariloche, Provincia
Rió Negro.
Imagen
ilustrativa. (*)
Eoceno. El fósil de la flor
margarita más antiguo del mundo, que data de unos 47 millones de años,
fue hallado cerca de la turística ciudad argentina de Bariloche. También se hallaron
restos de granos de polen. No hay registro de que supere su antigüedad
hasta ahora en el mundo. El hallazgo se hizo en 2008 y, después de las
investigaciones de laboratorio que determinaron su origen y antigüedad,
corroboradas por un equipo de científicos argentinos y suecos, fue
publicado en la más reciente edición de la prestigiosa revista
científica Science.
Solanaceae.
Fósil
de Solanaceae de la Patagonia Argentina.
Aspecto de Solanaceae
(*).
Eoceno.Son una familia de plantas herbáceas o leñosas con las hojas alternas,
simples y sin estípulas pertenecientes al orden Solanales, de las
dicotiledóneas. Las hojas son
generalmente alternas o alternadas a opuestas (o sea, alternas en la
base de la planta y opuestas hacia la inflorescencia). La consistencia
de las hojas puede ser herbácea, coriácea, o pueden estar transformadas
en espinas. En general las hojas son pecioladas o subsésiles, raramente
sésiles. La historia del origen de esta familia es en gran parte
desconocida ya que hasta ahora, solamente se habían encontrado algunas
semillas. Los científicos dicen que los orígenes de esta clase de fruto
se remontan decenas de millones de años, mucho más de lo que se pensaba
anteriormente. La planta, un tipo de Physalis, se encontró en una
selva fosilizada en la Patagonia.
Ceratopetalum edgardoromeroi.
Gandolfo, M. y Hermsen, E. 2017.
Ceratopetallum edgardoromeroi, del Eoceno
de Chubut. Prensa.
Aspecto de
Ceratopetallum
(*).
Eoceno.Recientemente acaba de
darse a conocer los resultados de un estudio realizado por
paleobotánicas de Argentina y Estados Unidos, sobre el fósil de una flor
madura (en realidad un fruto seco) que, con sus cinco tépalos leñosos a
modo de hélice, habría tenido una estrategia similar para dispersar sus
semillas. Estos fósiles tienen una antigüedad de 50 millones de años, y
fueron hallados en el noroeste de Chubut. El fósil corresponde a la
familia de las Cunoniáceas, cuya distribución actual incluye Sudamérica
y otras regiones distantes del Hemisferio Sur. Algunos representantes
actuales de esta familia en Patagonia incluyen la Tiaca y el Tineo que
crecen en zonas muy húmedas a ambos lados de la Cordillera de los Andes.
Ginkgoites patagonica.
Villar de Seoane et al. 2015.
Ejemplar recuperado en Chubut. MEF.
Eoceno.Están representados en la actualidad por una sola especie, Ginkgo biloba, ampliamente cultivada en todo el mundo.
Son y fueron vegetales arbóreos con hojas características por su
forma como abanico y con las venas divididas en horqueta
(dicotómicas). Son dioicas, es decir órganos sexuales están en
individuos separados, los poliníferos en los masculinos y los
ovulíferos en los femeninos. Los óvulos son, en la especie actual,
simples pedunculados. Su historia geológica se remonta a la era
paleozoica con el hallazgo de diferentes tipos foliares similares a
los actuales o mesozoicos, también referidos a la clase. En el
Eoceno de la Patagonia, se han encontrado importantes restos de este
grupo, con especimenes fértiles, portadores de numerosos óvulos
dispuestos en forma compacta (tipo estróbilo) y que se describieron
como género Karkenia. Es interesante destacar que en
Patagonia encontramos también formas paleozoicas con
fructificaciones que han sido referidas al grupo como posible stock
ancestral de los linajes mesozoicos.
Papuacedrus
prechilensis.
Peter Wilf
et al, 2009.
Fósil de
Papuacedrus prechilensis del Eoceno
de Río Pichileufú, en la provincia de
Rió Negro. Colección MEF.
Posible
aspecto en vida. (*):
Eoceno.Investigadoresestadounidenses y
argentinos han descubierto y analizado abundantes especímenes
fosilizados de una conífera conocida anteriormente como
Libocedrus prechilensis y encontrados en la Patagonia
argentina. Este vegetal fue descrito por primera vez en 1938, sobre
la base de un fósil cuyas características parecían coincidir más
estrechamente con las de una conífera actual cuyo nombre científico
es Austrocedrus (Libocedrus) chilensis. Sin embargo,
numerosas características de las hojas, incluyendo su forma
distintiva y la configuración de sus estomas, así como otros
detalles de los nuevos especímenes descubiertos, encajan a la
perfección con las típicas del actual género Papuacedrus,
estrechamente emparentado, y que hoy en día sólo está presente en
Nueva Guinea y las Islas Molucas. Basándose en los especímenes
fósiles recién descubiertos, de hace 52 y 47 millones de años, se
han reclasificado la especie fósil, catalogándola dentro del género
Papuacedrus, bajo el nuevo nombre de Papuacedrus
prechilensis. Una de las deducciones más importantes que los
científicos han hecho a raíz de esta nueva catalogación es que,
puesto que el género Papuacedrus suele hallarse en
hábitats tropicales montañosos y está limitado fisiológicamente a
los climas sumamente húmedos, en el Eoceno la Patagonia tuvo que ser
un lugar tropical cálido y húmedo, como ya han sugerido otros
indicios encontrados en estudios anteriores, y no una estepa fría y
seca como lo es hoy en su mayor parte.
Castanopsis
sp.
D.
Don.
Fósil de
Castanopsis sp del Eoceno de Patagonia.
Aspecto de la
planta de la familia de las fagaceas.
Eoceno.En la Patagonia
Argentina, se halló el primer registro fósil de este género de
plantas de la familia de las fagaceas en el hemisferio sur. El
descubrimiento contó con la participación de un investigador del
CONICET y fue publicado en Science. Hace dos
décadas, un equipo de investigación conformado por geólogos y
paleontólogos del CONICET en el Museo Egidio Feruglio y de la Pennsylvania
State University, iniciaron un proyecto conjunto con el objetivo de
poder estudiar y poner en valor el sitio de Laguna del Hunco, un
yacimiento de plantas fósiles de unos 52 millones de años (Eoceno
temprano) ubicado al noroeste de la provincia del Chubut, que si
bien era conocido desde los años ’20 del siglo pasado, hasta el
momento no había sido todavía suficientemente explorado. Durante los
últimos 20 años, este equipo de investigadores dio a conocer una
serie hallazgos de valiosos materiales fósiles (fundamentalmente de
vegetales) que los condujo a establecer la hipótesis de que la
descendencia de la flora que vivía en Patagonia hace 52 millones de
años, sobrevive hoy en día en los bosques de sudeste asiático y el
noreste de Australia, particularmente en la región biogeográfica
conocida como Australasia. Durante el
Eoceno, estas dos regiones, hoy distantes, se encontraban unidas a
través del continente Antártico (millones de años antes de que se
cubriera de hielo), que pudo oficiar de puente continental para el
flujo de plantas y animales entre ambas áreas. Si bien el cambio
drástico del clima que experimentó la Patagonia, que en ese momento
era tropical o subtropical, fundamentalmente como consecuencia del
ascenso de la Cordillera de los Andes y la separación de Sudamérica
de la península Antártica -que llevó a que las corrientes frías
provenientes de la Antártida ascendieran por el Atlántico Sur- hizo
que muchos grupos de plantas y animales desaparecieran, en la zona
australásica pudieron sobrevivir debido a que las condiciones
climáticas se mantuvieron constantes durante todo este tiempo. El hallazgo
reciente de restos fósiles de plantas del género Castanopsis (de la
familia de las fagaceas) en Laguna del Hunco, permite reconfirmar una
vez más la hipótesis de que antiguamente existió un tránsito
migratorio de especies vegetales entre la Patagonia y Australasia.
Pindocarpon
chichinalensis.
Muci, Passalia, e Iglesias. 2026.
Frutos fósiles de
Pindocarpon chichinalensis.
Aspecto de Pindocarpon
chichinalensis, mientras
un perezoso trepa la palmera en la antigua selva patagonica.
Mioceno. La gran abundancia de
frutos y semillas fósiles recuperados, que alcanzan cientos e
incluso miles de ejemplares distribuidos en distintos niveles
sedimentarios, indica que durante el Mioceno temprano existieron
extensos
palmares en esta región patagónica. Estos bosques
probablemente formaban parte de un paisaje abierto semejante a las
sabanas subtropicales modernas, donde las palmeras constituían
pequeños bosques intercalados con áreas de vegetación más baja. El
yacimiento de Paso Córdoba conserva una de las paleofloras más
importantes del Mioceno de Sudamérica, junto con abundantes fósiles
de mamíferos, aves, reptiles y plantas. La presencia de
Pindocarpon chichinalensisconstituye el primer
registro fósil inequívoco de la subtribu Attaleinae en el sur de
Sudamérica y uno de los frutos fósiles de palmera mejor preservados
conocidos a nivel mundial. Su descubrimiento aporta información
fundamental sobre la evolución de las palmeras, la historia de las
floras sudamericanas y las condiciones climáticas que dominaron el
norte de la Patagonia durante el Cenozoico temprano.
Paraalbizioxylon sp. Schuster, 1910.
Algunas de los numerosas
maderas fósiles halladas en la localidad de Termas de Rió
Hondo, atribuidas al Plioceno temprano. Colección del Museo
Municipal de Paleoantropología "Rincón de Atacama".
Mio-plioceno.
La familia Leguminosae o
Fabaceae se distribuye actualmente en
todos los continentes a excepción de la Antártida. Comprende
aproximadamente 720 géneros y con unas 18000 especies es la tercer
familia de angiospermas en cuanto a su número de representantes.
Es
el primer registro que se encuentra en la provincia de Santiago del
Estero, a través de estos materiales se pudo obtener sobre el
contexto ambiental de la época en leños fosilizados que fueron
hallados en la formación Choya (Mio-Plioceno) y es el primer
registro fósil de maderas de leguminosas. Estos hallazgos fueron
localizados a orillas del río Dulce en cercanías de la ciudad de
Termas de Rió Hondo, recuperados por Sebastián Sabater y pertenecen
a la colección del Museo Municipal de Municipal Rincón de
Atacama.
Rizolitos.Klappa, 1980.
Rizolitos (molde vegetales)
del Plioceno de Miramar. Colección Museo Municipal Punta Hermengo.
Rama mandibular de un
camelido gigante (Pleistoceno) con rizolitos sobre las
mismas. Colección Museo Municipal Punta Hermengo.
Plio-Pleistoceno.
El termino rizolito (tambien llamadas rizoconcreciones) describe aquella roca cuyos
razgos estructurales, texturales y origen son el resultado total o
mayoritariamente de la actividad de una antigua planta. Estas trazas de raíces
son de
variable morfología y concentraciónes. Los rizolitos son estructuras de
bioturbación diagnósticas de diagénesis meteórica y exposición
subaérea, características de ambientes continentales.Es menos habitual el
hallazgo de restos fósiles de vegetales, ya que las plantas están
conformadas principalmente por celulosa, sustancia que se descompone
fácilmente en condiciones normales.
Alternaria sp.
Nees, 1816.
Espora de
hongo Alternaria sp, recuperada entre los huesos de
las patas de un Gliptodonte en la localidad de San Pedro.
Presencia
característica de Alternaria sp, sobre una hoja en la
actualidad.
Forma de
estudio (*).
Pleistoceno.
Es un hongo ascomiceto, esto es, del filo de las
Ascomycotas. Las diferentes especies de este género son uno de los
mayores patógenos de plantas, y en menor medida puede afectar a los
animales a partir de su ingesta.
Alternaríaes un género fúngico muy común, donde se incluyen numerosas
especies de hongos saprofitas, endofíticas y patógenas ampliamente
distribuidas en el suelo y la materia orgánica en descomposición.
Las especies del género Alternaría sintetizan más de 70
metabolitos secundarios tóxicos para las plantas (fitotoxinas),
algunos de los cuales afectan a los animales, por lo que se
consideran micotoxinas. El Grupo
Conservacionista de Fósiles, equipo del Museo Paleontológico de San
Pedro, junto al Centro de Ecología Aplicada del Litoral
(CONICET-UNNE), de Corrientes, lograron muestrear e identificar
granos de polen y esporas fósilesde
diferentes plantas y hongosque habitaron los alrededores de un pantano cerca de la ciudad de
San Pedro, provincia de Buenos Aires, hace unos 700.000 años. El
material fue recuperado entre las patas de restos oseos de
Gliptodontes. Los datos
arrojados por los fósiles y el análisis del sedimento donde se
encontraban, condujeron a los investigadores a comprender que
aquellos pesados animales habían muerto en un antiguo pantano o
humedal que existía en la zona y que actuaba como “trampa natural”
cuando los grandes herbívoros se acercaban en busca de agua o
comida. Allí se
lograron separar e identificar los diminutos granos de polen y
esporas fósilescorrespondientes a
diferentes familiasde angiospermas
(plantas con flores), esporas de hongos y restos de algas
microscópicas que estarían indicando que se trataba de un ambiente
de estepa formada por plantas herbáceas, de suelos arenosos,
relacionada con cuerpos de agua cercanos.
Seguimos analizando la
Paleobotánica.
La
paleontología estudia los organismos que habitaron la tierra en el
pasado geológico. Lo hace a partir del análisis de la más importante
y a veces única evidencia que poseemos de ellos: sus restos fósiles.
Abarca principalmente tres grandes ramas: la paleontología de
vertebrados (reptiles, mamíferos, aves), la paleontología de
invertebrados (moluscos, entre otros) y la paleobotánica. A la
última se refieren los artículos de este número, dedicados a algunos
de los grupos de plantas que vivieron en o en las cercanías de lo
que hoy es el territorio argentino entre unos 500 y unos 45 millones
de años (Ma) antes del presente.
La
vida en la Tierra, tal como la conocemos hoy, es el resultado de los
cambios producidos a lo largo del tiempo por la evolución, de los
cuales los fósiles proporcionan un excelente testimonio. La
abundancia y la diversidad del registro fósil ponen en evidencia que
solo una pequeña fracción de la diversidad de formas de vida
pretéritas está hoy presente en el planeta.
Por lo
general, cuando un organismo muere sobreviene su
descomposición. Sin embargo, en condiciones particulares,
una pequeña fracción de los seres vivos que habitaron la
Tierra terminó conservada como fósiles. En el caso de las
plantas, esos fósiles pueden ser improntas o huellas sobre
rocas, compresiones carbonosas, permineralizaciones (tejidos
parcialmente reemplazados por minerales) o petrificaciones
(mineralización total). Los tejidos y órganos tienen
diferentes potenciales de fosilización: las hojas
resistentes y más duras, maderas, frutos leñosos, semillas y
polen son fósiles frecuentes, mientras que flores y frutos
delicados son escasos.
Croquis esquemáticos de los cambios en la configuración de
continentes y mares entre hace unos 600Ma y el presente. Cada grupo
de tres imágenes muestra, de izquierda a derecha, la visión
ecuatorial de la Tierra, del polo norte y del polo sur. Los esquemas
correspondientes a hace 300Ma y 260Ma indican la masa continental a
la que se dio el nombre de Pangea rodeada del océano primigenio
llamado Pantalasa. Hace 150Ma, por efecto de la deriva de los
continentes, Pangea se había dividido en los supercontinentes
Laurasia al norte y Gondwana al sur, mientras que se podían
diferenciar dos partes de Pantalasa: los océanos Pacífico y Tethys,
al tiempo que se insinuaba el Atlántico. Los siguientes esquemas dan
cuenta de la fragmentación de los supercontinentes hasta llegar a la
situación actual. En blanco, las principales áreas cubiertas de
hielo.
Por
lo general los fósiles son fragmentos de un único órgano del
espécimen original (por ejemplo, un tronco, una hoja o una semilla)
o la combinación de dos o más estructuras (por ejemplo, una flor
conectada con la rama que la portaba). Los que se encuentran juntos
o próximos pueden pertenecer a un único individuo o a varios, de una
o de más especies. Reconstruir un organismo del pasado o establecer
cómo eran los integrantes de una especie extinguida y cómo esta
evolucionó son tareas semejantes a armar un rompecabezas de
numerosísimas piezas.
Plantas
Las
plantas son organismos que desempeñaron un papel fundamental en la
evolución de la vida terrestre. Proporcionan alimento y resguardo a
muchos otros seres vivos, producen oxígeno, regulan la humedad y
contribuyen a la estabilidad del clima.
El
estudio del registro fósil de plantas nos permite establecer cuándo
se originaron los grandes grupos de ellas (como las plantas con
flores), sus momentos de diversificación y, en algunos casos, su
momento de extinción. Además, proporciona información para
comprender la evolución en el tiempo, tanto de los ecosistemas como
del clima del planeta.
EL BOSQUE PETRIFICADO DE JARAMILLO
Seguramente a muchos lectores de Ciencia Hoy la mención de plantas
fósiles les evoque imágenes de bosques petrificados, que
técnicamente son yacimientos de maderas fosilizadas generalmente
compuestos por grandes troncos más numerosos fragmentos y astillas
diseminados por el suelo. Esos bosques existen en todo el mundo y
son bastante abundantes en la Argentina, particularmente en zonas
con poca vegetación actual.
El más
extenso y conocido del país es el del Monumento Natural
Bosques Petrificados, en Jaramillo, Santa Cruz (a veces
llamado Cerro Madre e Hija). Los fósiles datan del Jurásico,
de hace unos 170Ma, e incluyen maderas, conos y hojas de
araucarias y otras coníferas. También son conocidos el
Monumento Provincial Bosque Petrificado Sarmiento, en Chubut,
del Paleoceno, y el Parque Paleontológico Araucarias de
Darwin, del Triásico, en Mendoza.
Las
maderas fósiles de estos bosques se estudian cortándoles láminas de
unos 50 micrómetros de espesor que, vistas al trasluz con un
microscopio, permiten identificar los distintos tipos de células que
las forman, sus características y su disposición, que son propias de
cada especie de árbol. Así, las maderas fósiles de Jaramillo
indicaron que abundaban las araucarias, y las de la península
antártica revelaron que durante gran parte del Cenozoico había allí
bosques similares a los patagónicos actuales.
El tiempo geológico
Con
los inicios de la geología moderna hacia el fin del siglo XVIII,
empezaron los esfuerzos por establecer el marco de referencia
temporal de la historia de la Tierra y de la vida en ella, es decir,
de definir la escala del tiempo geológico.
En los
comienzos y prácticamente todo a lo largo del siglo XIX, los
geólogos definieron cronologías relativas, por las cuales ordenaban
las rocas de un lugar o región según su antigüedad relativa, pero no
podían datarlas en años. Para los primeros años del siglo XX fue
posible intentar una cronología absoluta en Ma, por ejemplo
recurriendo a los isótopos radiactivos de diversos elementos, entre
otros métodos. Los progresos realizados en la construcción de esa
cronología desembocaron en la tabla reproducida en la página 16, que
muestra las principales divisiones del tiempo geológico entre el
momento de origen de la Tierra, hace unos 4600Ma, y el presente.
El desplazamiento de los continentes
En esos 4600Ma, no solo cambió la vida en nuestro
planeta: también varió la configuración de los continentes y los
mares. En 1912 el geofísico alemán Alfred Wegener (1880-1930),
basándose en el hecho de que los bordes de los continentes actuales
parecían encajar unos con otros como las piezas de un rompecabezas,
postuló que habrían sido parte de un supercontinente que terminó
llamándose Pangea. Ciertas evidencias, como la similitud de eventos
glaciares y registros fósiles en partes del planeta que hoy se
encuentran muy distanciadas, en su momento podían explicarse por la
teoría de Wegener, pero se desconocían los mecanismos que habrían
producido las cambiantes configuraciones de continentes y mares,
hasta que en la década de 1950 la tectónica de placas proveyó de
fundamento científico a lo que hoy llamamos deriva continental.
Cambios del clima
También el
clima experimentó profundos cambios a lo largo del tiempo geológico,
con momentos de amplias glaciaciones como las registradas en el
Carbonífero (hace unos 300Ma) o en el Pleistoceno (entre 2,6 y 0,1Ma
antes del presente), seguidas por intervalos cálidos en que floras y
faunas tropicales avanzaron hacia las altas latitudes, y tierras hoy
cubiertas por hielos permanentes, como la Antártida, tenían una
importante cubierta vegetal. El gráfico presenta una estimación de
cómo varió la temperatura terrestre desde hace unos 70Ma.
Evolución de la vida
La vida en
la Tierra evolucionó en el marco de los cambios comentados de los
continentes, los mares y el clima. Las primeras modalidades de vida
aparecieron en los océanos hace unos 3500Ma en forma de bacterias y
otros organismos unicelulares. La aparición de la fotosíntesis en
las cianobacterias y las algas verdes dio lugar a la progresiva
oxigenación de la atmósfera, la formación de la capa de ozono y el
establecimiento de condiciones apropiadas para el surgimiento de
formas de vida más elaboradas. De estas, las briofitas constituyen
el linaje más antiguo de las plantas terrestres cuyos primeros
registros datan de fines del Ordovícico y principios del Silúrico de
la era paleozoica, hace unos 500Ma (ver artículo ‘Primeros pasos de
la vida fuera del agua’).
Las plantas
se adaptaron a vivir en tierra a partir del desarrollo de tejidos
especializados, capaces de transportar internamente agua y
nutrientes. A partir del Devónico, hace unos 420Ma, se produjo un
importante recambio florístico en el que aparecieron nuevas formas
de plantas con raíces, tallos y frondes. Algunos grupos, como las
licofitas, se expandieron durante el Carbonífero, hace entre unos
350 y 300Ma, y formaron pequeños bosques; de otros, como los
helechos, entre ellos los arborescentes (ver artículo ‘Helechos
arborescentes en la Antártida’), llegarían unos pocos grupos hasta
nuestros días.
Los primeros
grupos de plantas terrestres se reproducían en un medio acuático
mediante esporas formadas en órganos especiales llamados
esporangios.
Los
grupos más evolucionados se reproducen por medio de
semillas, que es una forma más exitosa de reproducción ya
que permite a la planta independizarse del agua para
multiplicarse. Se reconocen dos subgrupos de plantas con
semillas: las gimnospermas y las angiospermas, antes de cuya
aparición, sin embargo, el registro fósil reveló la
existencia de helechos con semillas o pteridospermas, hoy
extinguidos, que constituyen un paso evolutivo intermedio
entre los helechos y las gimnospermas. Entre las
pteridospermas del Pérmico (entre 299 y 252Ma antes del
presente) se destacan las del orden Glossopteridales.
Las
gimnospermas se habrían originado en el Carbonífero (entre hace 359
y 299Ma) y expandido en el Pérmico con varios grupos extinguidos.
Habrían dominado los ecosistemas mesozoicos (entre hace 252 y 66Ma)
con nuevos linajes, algunos desaparecidos, como la familia
Cheirolepidiaceae (ver artículo ‘Pararaucaria y la evolución de las
coníferas’) y otros que llegan a nuestros días como las cícadas (ver
artículo ‘Cícadas, fósiles vivientes del reino vegetal’) o las
araucarias.
Las
angiospermas, por su parte, aparecieron por primera vez de manera
cierta en el registro fósil en el Cretácico temprano, hace unos
130Ma (ver artículo ‘Cuando las primaveras empezaron a tener
flores’), y se diversificaron rápidamente en el Cretácico tardío,
hace unos 75Ma, para prácticamente dominar durante el Cenozoico (a
partir de hace 65Ma) todos los ecosistemas (ver artículos ‘La
evolución temprana de las asteráceas’ y ‘Casuarinas y eucaliptos,
los árboles perdidos de la Patagonia’).
La paleobotánica en la Argentina
La
disciplina tiene un desarrollo relativamente reciente en el país,
comparado con el de otras ramas de la paleontología. En el siglo XIX,
numerosos naturalistas europeos, como Félix de Azara (1746-1821),
Alcide d’Orbigny (1802-1857), Charles Darwin (1809-1882) y Hermann
Burmeister (1807-1892), entre otros, comentaron hallazgos de plantas
fósiles realizados por ellos o por otros, lo mismo que hicieron
hacia fin de ese siglo exploradores locales como Francisco P Moreno
(1852-1929), Ramón Lista (1856-1898) y Jorge Fontana (1846-1920).
En el
siglo XX se destacaron la labor del berlinés radicado en
Córdoba Federico Kurtz (1854-1921), de quien la Academia
Nacional de Ciencias publicó póstumamente su Atlas de
plantas fósiles de la República Argentina; el médico y
naturalista ítaloargentino Joaquín Frenguelli (1883-1958) y
el geólogo italiano Egidio Feruglio (1897-1954), entre
otros.
El
despegue de la paleobotánica en la Argentina tuvo lugar en
la segunda mitad del siglo XX con la labor de Alberto
Castellanos (1896-1968), discípulo de Kurtz, y de sus
numerosos alumnos, que se cuentan entre los autores de las
primeras tesis doctorales en la especialidad escritas en el
país en las décadas de 1940 y 1950, entre ellos Pedro
Stipanicic (1921-2008), Carlos A Menéndez (1921-1976), María
Bonetti (1923-2015) y Sergio Archangelsky.
Estos, junto
con el botánico y palinólogo Juan Carlos Gamerro (1923-2013),
sentaron las bases para el florecimiento actual de la disciplina en
la Argentina.
Articulo originalmente publicado en Ciencia
Hoy (Numero 159. Volumen 26) cienciahoy.org.ar
está dedicado a la paleobotánica. Los editores agradecen la
eficiente colaboración de Viviana D Barreda, investigadora
principal del Conicet en el Museo Argentino de Ciencias
Naturales, que actuó como editora invitada para su
preparación. En tal carácter, definió los temas a tratar,
propuso autores y árbitros, revisó los manuscritos y
participó en las tareas de edición. Roberto R Pujana la
asistió en esas tareas y se desempeñó como enlace con el
comité editorial. Quede constancia del reconocimiento de
este por la labor de ambos. El texto e imágenes fueron
adaptados para este sitio.
De quienes son los fósiles?
En Argentina, por
ley, todos los restos fósiles hallados en su territorio forman parte
integral del Patrimonio Cultural de la Nación.
La Ley 25743/03 de
“Protección del Patrimonio Arqueológico y Paleontológico” regula
toda actividad relacionada al patrimonio paleontológico y el Museo
Argentino de Ciencias Naturales es el organismo nacional competente
de protección del patrimonio paleontológico, constituyéndose en la
Autoridad de Aplicación Nacional
de la Ley 25743.
Son sus
competencias, entre otras, llevar el Registro Nacional de
Yacimientos, Colecciones y Restos Paleontológicos, autorizar
el traslado de materiales en préstamo al exterior y
gestionar la repatriación de materiales paleontológicos
argentinos en el exterior.
Por otra
parte, las provincias argentinas, por la reforma de la
Constitución Nacional de 1995, son los custodios de los
materiales paleontológicos hallados en sus territorios y
encargadas de autorizar exploraciones, extracciones,
préstamos y traslados de estos materiales dentro del
territorio nacional.
<<<Los fósiles deben estar en los museos,
para el estudio y conocimiento de los visitantes y para las
futuras generaciones.
Es decir, que, si
encontrar un resto fósil (al menos en Argentina) debes comunicarlo
al museo o institución más cercana y competente. Los fósiles deben
estar en los museos como lo marca la ley. De esta forma ayudamos a
proteger nuestro patrimonio y hacemos posible que los investigadores
puedan estudiarlos. Si no vivís en Argentina y su país no tiuene una
legislación clara, también te invitamos a que te acerques a un museo
para contribuir con el patrimonio de tu nación.
Cícadas, fósiles vivientes
del reino vegetal.
Por Leandro CA.
Martínez y Analía E. Artabe. Publicado originalmente en Ciencia Hoy
Volumen 26 Número 154.
La historia de un grupo de plantas
presente en la Tierra desde hace casi 300Ma, por lo que podemos ver
a sus integrantes actuales como fósiles vivientes.
Las cícadas son un antiguo y
enigmático grupo de plantas con semillas, la mayoría de cuyas
especies tiene apariencia similar a las palmeras, con un tronco en
forma de columna, sin ramificar y por lo común recubierto de hojas,
que remata en una corona o penacho de hojas con sus extremos
divididos. También hay especies con tallo subterráneo, pero siempre
presentan el penacho de hojas en su ápice. Son comunes en plazas y
jardines, tanto por razones ornamentales como por su resistencia a
plagas y otros factores.
Aunque por su aspecto general
parezcan palmeras, las cícadas son gimnospermas, es decir, plantas
cuyas semillas no se forman en un ovario cerrado ni son parte de un
fruto, sino que están expuestas. Otras gimnospermas vivientes son
las coníferas (pinos, cipreses, araucarias, abetos, etcétera), las
gnetales (de las que existen hoy tres géneros, Gnetum, Ephedra y
Welwitschia, con unas setenta especies) y el caso único del Ginkgo
biloba o árbol de los 40 escudos.
Las cícadas pertenecen al linaje
más antiguo de plantas con semilla que vive en el presente: los
primeros registros de plantas del grupo datan de hace unos 290Ma, es
decir, del período pérmico, el último de la era paleozoica.
Las cícadas fueron diversas y muy abundantes
en los bosques y las selvas de la era mesozoica, cuando
vivieron los dinosaurios. Por ello, los botánicos se
refieren al Mesozoico como la era de las cícadas. Hacia
finales del Cretácico y en el límite con el Paleoceno, hace
unos 65Ma, junto con numerosas otras especies, sufrieron un
proceso de extinción cuya consecuencia es que llegaron a la
actualidad unos 10 géneros y unas 180 especies, todos
pertenecientes al orden Cycadales, compuesto por tres
familias: Cycadaceae, Stangeriaceae y Zamiaceae.
Son plantas que viven hoy en
lugares cálidos del planeta, en las Américas, el sudeste asiático,
Australia y África.
La mencionada extinción masiva del
Cretácico-Paleoceno no fue la única a la que sobrevivieron las
cícadas; antes de ella pasaron por otras dos: la del
Pérmico-Triásico hace unos 252Ma, y la del Triásico-Jurásico hace
unos 201Ma. Su éxito y su supervivencia hasta la actualidad quizá se
deba a que poseen adaptaciones para soportar sequía, fuego,
patógenos y predadores. Así, acumulan en sus tallos gran cantidad de
una sustancia viscosa llamada mucílago, que retiene agua
aprovechable en períodos de sequía. También sintetizan y acumulan
gran variedad de compuestos que les confieren resistencia a
patógenos o que alejan predadores por su carácter tóxico. De hecho,
la mayoría de los animales evitan comerlas.
Los estudios de los fósiles de
cícadas han aportado diversos datos de tipo biogeográfico (o sea, su
distribución en los continentes), paleoclimático y paleoecológico
(es decir, sobre las condiciones ambientales del pasado). La
información obtenida por medio de los estudios anatómicos y
morfológicos permiten establecer la relaciones filogenéticas (o de
parentesco) y entender tanto los procesos como las tendencias
evolutivas en este grupo de plantas.
Se han concebido diferentes
hipótesis sobre el origen evolutivo de las cícadas, definidas sobre
la base de dichos estudios anatómicos de fósiles. Entre ellas, se
destaca la que sostiene su descendencia de las pteridospermas o
helechos con semillas, que existieron a lo largo de las eras
paleozoica y mesozoica y eran un grupo muy diverso de plantas, con
numerosos órdenes y familias.
Más concretamente, dicha hipótesis
considera como probables antepasados de las cícadas a uno de los
órdenes de pteridospermas paleozoicas, el de las Medullosales, que
habitaron los bosques tropicales de hace unos 350Ma y eran plantas
con tallo en forma de columna, semejantes a una palmera pequeña o
también a lianas, con grandes hojas compuestas y semillas de
considerable tamaño. Estas características de su forma y apariencia,
además de otros análisis más complejos, de tipo anatómico y
filogenético, llevaron a postular el parentesco indicado.
Desde hace bastante, los
paleobotánicos piensan que el orden Cycadales es monofilético, lo
que significa que todos sus integrantes descienden de un ancestro
común. Llegaron a esa conclusión sobre la base de comparaciones
anatómicas y morfológicas entre los fósiles y las plantas actuales
que se han ido refinando en las últimas décadas. Para fósiles
vegetales, solo en contados casos se puede recurrir a los modernos
análisis de parentesco que permite la actual genética molecular,
porque salvo rara excepción no se encuentran en ellos restos de ADN.
A partir del inicio del período triásico,
hace unos 252Ma, cuando comienza la era mesozoica, se
registra la presencia de cícadas en diversos yacimientos
fosilíferos de todo el planeta. La Argentina no es la
excepción: en la cuenca geológica cuyana, en Mendoza, San
Juan y San Luis, se han encontrado hojas de cícadas
primitivas del género Pseudoctenis; y en los sedimentos de
Ischigualasto, en San Juan, se halló un tallo fosilizado del
género Michelilloa, cuyas características sugieren que es un
temprano integrante del orden Cycadales.
Los fósiles
encontrados en diversas partes del mundo permiten concluir que el
número de géneros de Cycadales se incrementó a lo largo del
Mesozoico, y que el orden alcanzó una gran diversidad en el
Cretácico, como se aprecia por la elevada variedad de hojas,
semillas y troncos fósiles encontrados. En los yacimientos del
Mesozoico de la Argentina se identificaron varios géneros fósiles de
hojas y leños, además de numerosas especies clasificadas como afines
con las Cycadales.
Con la extinción
masiva de especies de fines del período cretácico, que acabó con las
pteridospermas (lo mismo que con los dinosaurios), desapareció la
mayoría de los géneros de Cycadales, pero el orden no se extinguió
por completo.
Por otro lado,
durante el transcurso del Cretácico aparecieron las primeras
angiospermas o plantas con flores, que se fueron diversificado y,
para el fin del período, habían colonizado y llegado a dominar los
diferentes ecosistemas terrestres. Ese dominio se incrementó luego
de la extinción del Cretácico-Paleoceno, pues las angiospermas
sobrevivientes terminaron ocupando los nichos ecológicos dejados por
otras plantas extinguidas, y así produjeron una transformación de la
apariencia, composición y ecología en la mayoría de los ecosistemas
del planeta.
En los yacimientos
fosilíferos cretácicos de la Patagonia se han encontrado numerosos
restos del orden Cycadales, tanto tallos y hojas como estructuras
reproductivas y semillas. Su estudio puso de manifiesto el
incremento y la diversificación de un grupo o subfamilia de la
familia Zamiaceae conocido con el nombre de Encephalarteae, la que
en la actualidad vive en las regiones tropicales y subtropicales de
África y Australia y agrupa tres géneros: Encephalartos en África, y
Macrozamia y Lepidozamia en Australia.
Muchos de los
fósiles cretácicos de Cycadales encontrados en la Patagonia fueron
asignados a la subfamilia Encephalarteae. Esta asignación es
coherente con el hecho de que durante la mayor parte del Mesozoico,
América del Sur, África, Australia, la India, Madagascar y la
Antártida conformaron parte del supercontinente Gondwana. Se puede
pensar que las Encephalarteae se diversificaron a lo largo de
Gondwana mientras perduró un clima templado-cálido. Luego, con el
cambio climático de fines del Cretácico y con la fragmentación de
Gondwana, se produjo por un lado la disyunción de las que estaban en
África y Australia y, por otro, la extinción de las existentes en lo
que hoy es la Patagonia.
Los cambios que se
produjeron luego del límite Cretácico-Paleoceno (o
Mesozoico-Cenozoico) incluyeron el levantamiento de la cordillera de
los Andes, el desplazamiento de la Antártida hacia el polo sur, la
formación de la corriente circumpolar antártica y la disminución de
los niveles de dióxido de carbono en la atmósfera. El resultado
final para la Patagonia fue a grandes rasgos su paso de un clima
templado-cálido a uno templado-frío, condiciones que se fueron
intensificando hasta el presente.
Algunas Cycadales
extinguidas, de los géneros Menucoa y Bororoa, fueron
halladas en sedimentos patagónicos de hace 63Ma, es decir
del Paleoceno, lo que demuestra que su extinción por los
factores mencionados tuvo una relativa gradualidad, lo mismo
que el pasaje de las regiones australes de Sudamérica del
clima cálido y húmedo a frío y seco que finalmente condujo a
la desaparición de las comunidades de bosques
templado-cálidos de la Patagonia y la Antártida, con
numerosos grupos de animales que vivían en ellas.
No se han hallado
en territorio argentino restos de Cycadales en estratos rocosos
posteriores al Eoceno, y hoy solo vive un género de ese orden en
Sudamérica, Zamia, con una distribución que va desde Colombia hasta
Bolivia. Está emparentado con el mencionado género fósil Bororoa
encontrado en estratos del Paleoceno en la Patagonia.
Las cícadas
cohabitaron con los dinosaurios y otros grupos de animales a lo
largo de la era mesozoica. Algunas de sus características indicarían
una posible coevolución con ciertos grupos de ellos. Las cícadas
poseen una colorida cubierta carnosa que envuelve sus semillas y es
consumida actualmente por numerosos animales (roedores, elefantes,
emúes, canguros, zarigüeyas, aves). Esto sugiere que en el pasado
podría haber sido consumida –y de este modo dispersada– por
dinosaurios, entre otros. Como en los procesos de extinción de
finales del Cretácico habrían existido efectos recíprocos entre
plantas y animales, la desaparición de los dinosaurios podría haber
producido ciertas alteraciones en las formas de dispersión de las
cícadas y limitado su migración. En otras palabras, el no poder
dispersarse o migrar hacia regiones más cálidas, sumado al
enfriamiento del clima, quizá haya sido uno de los factores que
condujo a la extinción de estas plantas en la Patagonia y en otras
regiones del mundo.
A pesar de haber
quedado reducido a un número pequeño de especies en comparación con
las que existieron en el remoto pasado geológico, las cícadas pueden
considerarse un grupo exitoso, ya que han permanecido en la Tierra
por cerca de 300Ma. Sin embargo, las especies actuales corren hoy el
riesgo de desaparecer por la destrucción de los ambientes en los que
habitan.
Lecturas Sugeridas
ARTABE AE & STEVENSON DW, 1999,
‘Fossil Cycadales of Argentina’, The Botanical Review, 65: 219-238.
CÚNEO NR et al., 2010, ‘Review
of the Cycads and Bennettitaleans from the Mesozoic of Argentina’,
en Gee CT (ed.), Plants in Mesozoic Time: Morphological innovations,
phylogeny, ecosystems, Indiana University Press, pp. 187-212.
MARTÍNEZ LCA, ARTABE AE & BODNAR
J, 2012, ‘A new cycad stem from the Cretaceous in Argentina and its
phylogenetic relationships with other Cycadales’, Botanical Journal
of the Linnean Society, 170: 436-458.
NORSTOG K & NICHOLLS T, 1997,
The Biology of the Cycads, Cornell University Press, Ithaca NY.
PANT DD, 2002, An Introduction
to Gymnosperms, Cycas and Cycadales, Birbal Sahni Institute of
Palaeobotany, Lucknow, India.
STEVENSON DW, 1990, ‘Morphology
and systematics of the Cycadales’, en Stevenson DW (ed.), The
Biology, Structure and Systematics of the Cycadaen, New York
Botanical Botanical Garden, 57: 8-55.
Helechos arborescentes
en la Antártida.
Por Lecturas
Sugeridas y Silvia N Césari. Publicado originalmente en la
Revista Ciencia Hoy. Volumen 26. Número 154. Adaptada para
Paleo.
La historia de los helechos
arborescentes de la Antártida, componentes abundantes y diversos
de la vegetación de hace 120 millones de años.
Cuando pensamos en helechos, la
primera imagen que nos viene a la mente es la de una planta pequeña,
con hojas (llamadas frondes) divididas en segmentos, que requiere
abundante agua, se reproduce mediante esporas y forma parte de la
decoración de interiores. Esa imagen coincide con muchas de las más
de diez mil especies de helechos que actualmente pueblan la Tierra,
pero existen algunas que escapan a tal descripción, como las del
orden Cyatheales.
Los helechos de
estas especies alcanzan alturas comparables con las de los árboles,
lo cual les valió el nombre de helechos arborescentes. Viven por lo
general en lugares cálidos y húmedos, como muchas zonas de Misiones,
Corrientes, Jujuy y Salta en la Argentina. Por su aspecto se pueden
confundir con palmeras, ya que presentan un eje vertical sin
ramificaciones, muchas veces con las bases de las hojas adheridas a
él, y una corona de hojas con forma de sombrilla en el extremo
superior.
Pueden superar los 15m de
alto, pero no son árboles sino helechos, como sus parientes
más pequeños y conocidos, pues se reproducen mediante las
esporas contenidas en pequeñas estructuras llamadas
esporangios, ubicadas en la parte inferior o envés de las
hojas. Las especies vivientes de Cyatheales conforman ocho
familias (Thyrsopteridaceae, Loxsomataceae, Culcitaceae,
Pagiogyriaceae, Cyatheaceae, Cibotiaceae, Dicksoniaceae y
Metaxyaceae).
En adición a
ellas, existen registros fósiles que sugieren que este grupo se
habría originado hace más de 150Ma, en el período jurásico.
Estudiando restos fósiles se han identificado numerosas especies
extinguidas, algunas de las cuales presentan caracteres que las
relacionan con familias actuales, pero otras tienen características
únicas, o incluso combinaciones de características propias de dos o
más familias poco emparentadas, por lo que su clasificación sobre la
base de los esquemas actuales resulta difícil. Esto revela que en el
pasado la diversidad del grupo fue mayor que la de hoy.
Fósiles bajo el
hielo
En los últimos
años, el grupo de investigación que integran los autores ha
descripto numerosos Cyatheales fósiles hallados en la península
Byers, en el extremo oeste de la isla Livingston, una de las
Shetland del Sur. Los hallazgos fueron realizados por expediciones
del Instituto Antártico Argentino y provienen de afloramientos
rocosos de la formación geológica Cerro Negro, de 120 millones de
años de antigüedad, es decir, del Cretácico inferior. Son rocas
presuntamente formadas por la acción de ríos y lagos ubicados en las
cercanías de volcanes, cuyas erupciones habrían aportado grandes
cantidades de cenizas y otros productos que se acumularon en forma
de sedimentos.
Los fósiles de la
península Byers demuestran que existió en la región una flora con
abundancia de coníferas, helechos, cícadas y otros grupos
extinguidos de gimnospermas, como las Bennettitales y las
Pteridospermas.
Sobre la base de los frondes
fósiles, se ha logrado distinguir tres especies del orden
Cyatheales, denominadas Eocyathea remesaliae, Sergioa
austrina y Lophosoria cupulata. No solo se advierten en
muchos fósiles detalles de las pequeñas venas de los frondes
sino, también, parte de sus estructuras reproductivas. Las
esporas generadas en los esporangios permanecen en ellos
hasta que maduran y estos se abren, lo que permite la
dispersión de las esporas.
Los esporangios de
los helechos pueden presentarse aislados, o en agrupaciones
denominadas soros o sinangios. El hallazgo de fósiles de helechos
con esporangios y esporas preservadas es importante porque permite
establecer con más certeza las afinidades del fósil. Muchos grupos
de helechos no emparentados entre ellos (e incluso otros grupos de
plantas que no son helechos) pueden tener frondes muy similares, en
cuyos casos el hallazgo de estructuras reproductivas resulta clave
para poder clasificarlos de manera precisa.
Además de
impresiones de hojas, se hallaron en la península Byers tallos
petrificados de helechos Cyatheales. En total se estudiaron restos
de unos ochenta ejemplares pertenecientes a cuatro especies
extinguidas de estos helechos. Por las particularidades de su
preservación, se pueden seccionar con las herramientas usadas para
cortar rocas, las cuales permiten obtener láminas delgadas para ser
estudiadas usando lupa o microscopio. Esto revela la disposición de
los tejidos y, en algunos casos, detalles de las paredes celulares.
Y el estudio de su anatomía hace posible comprender las razones de
que los helechos arborescentes alcancen tamaños tan altos.
Los estudios sobre
la geología y la flora fósil de la península Byers llevan a pensar
que, en el lapso de uno o dos millones de años en que se formaron
los estratos rocosos que contienen la flora fósil de la isla
Livingston, sería posible distinguir dos etapas de sucesión
florística. No se ha recogido suficiente evidencia como para estimar
el tiempo en el que se ubicaría el límite entre ambas etapas.
En la primera
etapa, en ambientes fluviales con esporádicas caídas de ceniza
volcánica, dominaron los bosques de coníferas formados por
araucariáceas y podocarpáceas de más de 20m de altura, acompañadas
en menor abundancia por helechos y otras gimnospermas. En la segunda
etapa, debido a un incremento de la actividad volcánica, el ambiente
se volvió desfavorable para las coníferas y la flora evolucionó
hacia una asociación de Cyatheales, Bennettitales –que son un grupo
de gimnospermas extinguidas que poseen hojas parecidas a las de las
Cycas–, helechos de pequeño porte y otras gimnospermas no coníferas,
con escasa presencia de araucariáceas y podocarpáceas.
Los fósiles de helechos
Cyatheales encontrados en la península Byers provienen de
rocas formadas en la segunda de las anteriores etapas. En
ella, períodos de mayor adversidad relativa del medio
habrían afectado a todos los grupos de plantas, pero los
helechos habrían tenido la capacidad de recuperarse y crecer
más rápido que las plantas que producen madera. Por ello,
sus formas arborescentes habrían podido alcanzar alturas
importantes con más facilidad que las gimnospermas.
Se ha estimado,
mediante modelos matemáticos basados en datos de plantas actuales,
que algunos de los Cyatheales de la península Byers habrían
alcanzado los 20m de alto. Si bien no lograron las alturas de hasta
45m estimadas para las coníferas de esa flora fósil, por su
abundancia y porte estimado fueron un elemento importante del
estrato arbóreo de dicha segunda etapa, en la que habrían tenido una
presencia mucho más importante que las coníferas.
Los restos
paleoflorísticos constituyen una de las herramientas usadas para
estimar cómo fue el clima durante el pasado geológico. Dado que las
plantas subsisten en los lugares en que las condiciones del medio
les son favorables, puede concluirse que la abundancia de
determinadas especies en una época y un lugar es señal de que allí y
entonces el medio ofrecía condiciones propicias para su
subsistencia.
Algunas de las
técnicas más difundidas para estimar los climas del pasado se
aplican a las angiospermas dicotiledóneas que, si bien ya habían
aparecido en la Tierra hace 120Ma, aún estaban en etapas tempranas
de su desarrollo y diversificación, y todavía no habían colonizado
varias regiones del planeta, entre ellas las tierras que hoy son la
isla Livingston.
Una técnica
alternativa, llamada ‘análogo moderno’, consiste en determinar qué
características posee el ambiente en que viven los representantes
actuales del grupo de plantas fósiles en cuestión y extrapolar a
estas esos requerimientos ambientales.
Los helechos
requieren que haya agua en el ambiente, ya que su reproducción
depende en parte de que los gametos masculinos se desplacen por ella
y lleguen a los femeninos. Ello explica que, por lo general, crezcan
en ambientes húmedos, los que muchas veces son también templados o
cálidos. El hallazgo de gran cantidad de helechos fósiles suele
tomarse como indicador de que imperó ese tipo de condiciones y, como
los Cyatheales actuales suelen vivir en ese tipo de sitios, su
hallazgo en gran abundancia y diversidad en la paleoflora de la isla
Livingston permite inferir que el clima les debería haber sido allí
bastante favorable durante el Cretácico y que imperaron similares
condiciones ambientales a aquellas en que prosperan los helechos de
hoy.
El hallazgo en la
isla Livingston de la especie fósil Lophosoria cupulata, una
dicksoniácea muy similar a la actual Lophosoria quadripinnata que
habita desde México hasta la Patagonia, permitió estimar que hace
120Ma la temperatura media anual en esa región de la Antártida
habría sido como mínimo de 8ºC. Por otro lado, la presencia de
numerosos grupos de helechos preservados allí –tanto de la familia
Cyatheales como de otras– llevó a postular que posiblemente no
existieron períodos de suelos congelados.
Un estudio de las
condiciones climáticas en que vivieron los Cyatheales y otros
helechos arborescentes actuales concluyó que el ancestro común de un
subconjunto de los primeros, que informalmente se denomina ‘núcleo
de los helechos arborescentes’, habría requerido un régimen alto de
precipitaciones, de por lo menos 1100mm anuales, con temperaturas
mínimas anuales no menores de 7,7ºC, y temperaturas máximas anuales
no menores de 23,3ºC. Dado que la mayoría de los helechos fósiles
del orden Cyatheales de la isla Livingston habrían
sido parte de
dicho núcleo, se puede concluir que el régimen de lluvias y las
temperaturas habrían configurado un clima muy benigno, bien distinto
del que hoy impera en la región.
Lecturas Sugeridas
CÉSARI SN, 2006,
‘Aptian ferns with in situ spores from the South Shetland Islands’,
Review of Palaeobotany and Palynology, 138: 227-238.
CÉSARI
SN, REMESAL M y PARICA C, 2001, ‘Ferns: A palaeoclimatic significant
component of the Cretaceous flora from Livingston Island,
Antarctica’, en Asociación Paleontológica Argentina, Publicación
especial 7, pp. 45-50.
VERA EI, 2013, ‘New
Cyathealean tree fern Yavanna chimaerica from the Early Cretaceous
of Livingston Island, Antarctica’, Cretaceous Research, 44: 214-222.
VERA EI, 2015,
‘Further evidence supporting high diversity of Cyathealean tree
ferns in the Early Cretaceous of Antarctica’, Cretaceous Research,
56: 141-154
Casuarinas y eucaliptos, los
árboles perdidos de la Patagonia.
Por María del
Carmen Zamaloa (Doctora en biología, UBA. Integrante del
Departamento de Ecología, Genética y Evolución, FCEN, UBA) y María
Alejandra Gandolfo (Doctora en biología, UBA. Investigadora del LH
Bailey Hortorium, Universidad de Cornell). Publicado originalmente
en Volumen 26. Número 154.
Dos familias de árboles que ahora
son naturales de Australia y lugares vecinos vivieron por unos 10Ma
en la Patagonia, cuando esta recibía unos 1100mm por año de lluvia y
registraba temperaturas medias anuales del orden de los 17˚C, e
invernales mayores que 10˚C.
Desde principios
del siglo XX aparecieron en la literatura especializada menciones
esporádicas de varios yacimientos de plantas fósiles de la era
cenozoica (iniciada hace 66Ma) en la Patagonia, pero solo en tiempos
recientes se comenzó a estudiar el tema en extensión y en
profundidad, tanto en materia de identificación de especies como de
dataciones y de colección controlada de ejemplares.
Paradigmático de
esos yacimientos es el de Laguna del Hunco, ubicado en la estepa
patagónica en el noroeste de Chubut, aproximadamente a mitad camino
en línea recta entre Esquel y Gastre. Allí, las rocas de unas
colinas bajas albergan restos fósiles de una rica paleobiota
compuesta por plantas, peces, ranas e insectos, en excelente estado
de preservación.
Es un excepcional
repositorio, uno de los de mayor diversidad florística cenozoica del
mundo, que permite conocer y comprender la dinámica de los antiguos
ecosistemas y su historia evolutiva.
De la gran diversidad
biológica encontrada en el citado yacimiento se destacan dos
grupos de plantas muy interesantes desde el punto de vista
fitogeográfico, ecológico y evolutivo: las casuarinas y los
eucaliptos. Ambos reúnen un par de condiciones muy valoradas
por los paleobotánicos: (i) tienen caracteres morfológicos
distintivos que facilitan su reconocimiento, y (ii) tienen
follaje, tallos y flores firmes, así como frutos leñosos, lo
cual les confiere alta resistencia a la descomposición y
aumenta sus posibilidades de convertirse en fósiles.
Las dataciones de
las rocas portadoras de los fósiles de Laguna del Hunco realizadas
por isótopos de argón permitieron establecer una antigüedad de 52Ma,
que corresponde al óptimo climático del Eoceno temprano, un período
en el que los climas mundiales eran más cálidos y húmedos que los
actuales. Se estima que lo que hoy es la Patagonia habría tenido
entonces las lluvias por encima de los 1100mm por año y temperaturas
medias anuales del orden de los 17˚C, con medias en invierno mayores
que 10˚C. La vegetación boscosa que crecía cerca de cuerpos
lacustres dominaba el paisaje.
Casuarinas
Las casuarináceas
son angiospermas (véase en este número el artículo ‘Cuando las
primaveras empezaron a tener flores. La historia evolutiva de las
angiospermas patagónicas’) que viven en climas subtropicales y que
en la actualidad solo crecen naturalmente en el sudeste asiático,
Australia y Melanesia. Son ampliamente cultivadas en zonas
subtropicales y templadas por su madera, para fijar suelos,
constituir barreras rompevientos y como plantas ornamentales. Sus
cuatro géneros (Casuarina, Allocasuarina, Gymnostoma y Ceuthostoma)
reúnen unas noventa especies y se destacan porque sus últimas ramas
son delgadas, verdes, estriadas y colgantes. Estas características
recuerdan las plumas de los casuarios, aves no voladoras de
Australia y Nueva Guinea, que son el origen de su nombre.
Las hojas de las
casuarinas se reducen a un anillo de dientes que apenas cubre los
nudos de las ramas y, a pesar de ser angiospermas, las plantas
carecen de flores vistosas: las femeninas son pequeñas y aparecen
agrupadas en conos que se vuelven leñosos al formarse los frutos,
mientras las masculinas, productoras de polen, forman espigas laxas
poco llamativas en los extremos de las ramas jóvenes. Estas
características recuerdan a las coníferas y, de hecho, en algunos
lugares se da a las casuarinas cultivadas el nombre de pino
australiano.
El primer
registro fósil de casuarinas en Sudamérica data de 1943 y se
debe al médico y naturalista italiano, radicado en Santa Fe
y profesor de las universidades nacionales del Litoral y La
Plata, Joaquín Frenguelli (1883-1958), quien halló en Laguna
del Hunco una rama con cuatro conos que denominó Casuarina
patagonica. Más tarde, sucesivos paleontólogos encontraron
en otras pocas localidades patagónicas granos de polen
dispersos en sedimentos de edades entre el Paleoceno y el
Eoceno.
Los hallazgos
realizados en Laguna del Hunco en los últimos quince años, por
varios grupos de investigación en los que participaron las autoras,
confirmaron ampliamente que existieron casuarináceas en la Patagonia,
y completaron un registro que incluye abundantes restos de follaje,
ramas con diferentes tipos de conos fructíferos que corresponden por
lo menos a tres especies y también ramas con pequeñas espigas de
flores que contienen granos de polen. Las posiciones de los fósiles
en los mismos estratos rocosos indican que varias especies de
casuarinas coexistían en la región.
Entre las
características morfológicas de esos fósiles se advierten ramas
estriadas, hojas reducidas a cuatro dientes en los nudos,
inflorescencias formadas por conos de aparente consistencia leñosa y
polen recuperado de espigas terminales. Ellas son las distintivas de
la familia, en particular, del género Gymnostoma.
Los fósiles
patagónicos son los únicos de casuarináceas hallados fuera de la
región australiano-asiática. Su diversidad y abundancia muestran que
las casuarinas eran un importante y bien establecido componente de
la flora patagónica a principios del Eoceno y señalan que la familia
tenía en ese tiempo una amplia distribución en Gondwana.
Eucalyptus es un
género de la familia de las mirtáceas que incluye unas 600 especies
endémicas de Australia e islas vecinas. Son componentes icónicos de
la flora australiana, y desempeñan un papel dominante en la
vegetación de esa zona del mundo tanto por la producción de biomasa
como por la diversidad de especies. Como plantas nativas ocupan un
amplio rango de ecosistemas, desde desiertos hasta selvas
tropicales, y como cultivadas se diseminaron a todo el mundo por el
crecimiento rápido de algunas de sus especies, por los usos
múltiples de su madera (papel, postes, construcciones rurales,
carbón de leña, cajonería, carpintería rústica, pisos), por los
aceites con propiedades medicinales (antisépticas, cicatrizantes,
antioxidantes) extraídos de sus hojas, como barreras rompevientos y
como plantas ornamentales.
Los eucaliptos se
caracterizan por tener hojas en forma de hoz con glándulas de aceite
que aparecen como pequeños puntos sobre la superficie, y flores
agrupadas en inflorescencias, siempre en número impar. Cada yema
floral o capullo tiene un capuchón que se desprende cuando la flor
se abre y deja expuestos los numerosos estambres (estructuras
productoras de polen) que son las partes más coloridas y llamativas
de la flor. Esto los diferencia de la mayoría de las plantas que
producen flores, cuyo principal atractivo es el perianto (cáliz y
corola). El fruto es una cápsula leñosa que se abre y libera las
semillas cuando madura.
También Frenguelli fue el
primero en describir un fósil de eucalipto en la Patagonia.
En 1953, publicó un trabajo titulado ‘Restos del género
Eucalyptus en el Mioceno del Neuquén’ (Notas del Museo de La
Plata, 16, 97: 209-213), en el cual se refirió a un fósil
compuesto por tres cápsulas hallado en rocas de edad
imprecisa entre el Oligoceno tardío y el Mioceno temprano
(28 a 17Ma).
Tras la
publicación de este fósil, el único hasta ese momento encontrado
fuera de la región australiano-asiática, casi nada se publicó sobre
posibles eucaliptos fósiles patagónicos hasta que en campañas
realizadas a partir de 1999 se descubrieron en Laguna del Hunco
abundantes improntas de hojas, frutos en grupos y solitarios,
capullos y flores con polen que al encontrarse asociados en los
mismos estratos de roca sugieren que proceden de la misma especie
biológica. Tomaron parte en esas campañas, financiadas en gran
medida por la National Science Foundation de los Estados Unidos,
investigadores del Museo Egidio Feruglio de Trelew, la UBA, la
Universidad de Cornell y Penn State University.
El mencionado
óptimo climático del Eoceno duró un lapso relativamente breve del
tiempo geológico, que parece coincidir aproximadamente con el
período del que datan los fósiles patagónicos de casuarinas y
eucaliptos, y fuera del cual no se han encontrado en Sudamérica
fósiles de ninguna de las dos familias. Su extinción posiblemente se
deba a los cambios ecológicos y ambientales producidos por el cambio
climático acaecido en el Eoceno medio, y cuyas consecuencias en las
floras fueron no solo extinciones de linajes completos, sino también
reducción de áreas geográficas de distribución, expansión de
especies mejor adaptadas a las nuevas condiciones, desplazamiento de
otras y aparición de nuevas especies.
El cambio de las
condiciones ambientales pudo haber sido menos abrupto en la región
de Australasia, lo que habría permitido la supervivencia de dichas
familias hasta la actualidad.
Lecturas Sugeridas
GANDOLFO MA et al.,
2011, ‘Oldest known Eucalyptus macrofossils are from South America’,
PLOS ONE, 6, 6: e21084. doi:10.1371/journal.pone.0021084.
HERMSEN EJ, GANDOLFO
MA & ZAMALOA MC, 2012, ‘The fossil record of Eucalyptus in Patagonia’,
American Journal of Botany, 99: 1356-1374.
WILF P et al., 2005,
‘Eocene plant diversity at Laguna del Hunco and río Pichileufu’, The
American Naturalist, 165: 634-650.
ZAMALOA MC et al.,
2006, ‘Casuarinaceae from the Eocene of Patagonia, Argentina’,
International Journal of Plant Sciences, 167: 1279-1289.
Bibliografía Sugerida.
Anzótegui, L. M. 1998. Hojas
de angiospermas de la Formación Palo Pintado, Mioceno
superior, Salta, Argentina. Parte I: Anacardiaceae,
Lauraceae y Moraceae. Ameghiniana, 35: 25-32.
Anzótegui, L.M. 2002. Hojas
de Myrtaceae en la Formación San José (Mioceno Medio)
Provincia de Tucumán, Argentina. Ameghinian, 39:
321-330
Anzótegui, L.M. 2004.
Megaflora de la Formación Chiquimil, (Mioceno Superior), en
los valles de Santa Maria y Villavil, provincias de
Catamarca y Tucumán, Argentina. Ameghiniana, 41: 303-
Anzótegui, L.M., Garralla, S.
y Herbst, R. 2007. Fabaceae de la Formación El Morterito,
(Mioceno Superior) del valle del Cajón, provincia de
Catamarca, Argentina. Ameghiniana, 44: 183-196
Anzótegui, L. M. y. Herbst,
R. 2004. Megaflora (hojas y frutos) de la Formación San José
(Mioceno Medio) en río Seco, departamento Santa María,
provincia de Catamarca, Argentina. Ameghiniana,
Azcuy, C.L., Laffitte, G. y
Rodrigo, L. 1984. El límite Carbónico-Pérmico en la cuenca
Tarija-Titicaca. 3º Congreso Argentino de Paleontología y
Bioestratigrafía, Corrientes, 1982, Actas 1: 39- 44.
Barreda, V.D, Anzótegui, L.M.,
Prieto, A.R., Aceñolaza, P., Bianchi, M.M., Borromei, A.M.,
BREA, M. Caccavari, M., Cuadrado, G.A., Garralla, S. Grill,
S., Guerstein, G.R., Lutz, A.I., Mancini, M.V., Mautino, L.R.,
Ottone, E.G., Quattrocchio, M.E., Romero, E.J., Zamaloa, M.C.
y Zucol, A. 2007. Diversificación y cambios de las
angiospermas durante el Neógeno en Argentina.Ameghiniana
50º Aniversario - Publicación Especial, 11: 173-191
de Benedetti, J.J.P. 1948.
Notas preliminares sobre el Devónico de la parte oriental de
la Sierra de Zapla y del Cerro Calilegua. Publicación
Instituto de Geología y Minería, Universidad Nacional de
Tucumán, 157:3
de la Puente, G.S. y Rubinstein, C.V. 2006. Primer hallazgo
de quitinozoos en el Tremadociano superior de la región de
Pascha-Incamayo, Cordillera Oriental, Argentina.
9º Congreso Argentino
Durango de Cabrera, J. ,
Georgieff, S.M. y Moreno, P. 1997. Primer registro del
género Equisetum Linneo (Pteridophyta) en la
Formación Andalhuala (Mioceno Superior-Plioceno), valle de
Santa María, Catamarca. Ameghiniana, 34: 119
Durango de Cabrera, J. y
Vergel, M.M. 1984. Primeros resultados del análisis del
paleomicroplancton de la perforación YPF St. LdC x-1
(Laguna del Cielo), Devónico de Salta. Reunión de
Comunicaciones A.P.A. Circular Informativa, 13: 4
Feruglio, E. 1931.
Observaciones geológicas en las provincias de Salta y Jujuy.
I Reunión Nacional Geografía, Contribuciones
Dirección General de YPF, 7: 5-39
Garcia, A. y Herbst, R. 1997.
Charophyta del Mioceno Superior de los valles de Santa María
y del Cajón, provincia de Catamarca, Argentina.
Ameghiniana, 34: 61-68
Gavriloff, I.C., Durango de
Cabrera, J. y Vergel, M. 1998. Paleontología de
invertebrados, paleobotánica y palinología. En M.
Gianfrancisco, M.E. Puchulu, J. Durango de Cabrera y G.F.
Aceñolaza (Eds.). Geología de Tucumán. Publicación
Especial Colegio Graduados Ciencias Geológicas Tucumán.,
211-226
Herbst, R., Anzótegui, L.M.,
Esteban, G., Mautino, L.R., Morton, L.S. y Nassif, N. 2000.
Síntesis paleontológica del Mioceno de los valles
Calchaquíes, noroeste argentino. En: F.G. Aceñolaza y R.
Herbst (Eds.) El Neógeno de Argentina. INSUGEO,
Serie Correlación Geológica, 14: 263-288
Herbst, R., Anzótegui, L.M. y
Jalfin, G. 1987. Estratigrafía, paleoambnientes y dos
especies de Salvinia Adanson (Filicopsida), del
Mioceno Superior de Salta, Argentina. Facena 7: 15-42
Lutz, A.I., 1987. Estudio
anatómico de maderas Terciarias del valle de Santa María
(Catamarca-Tucumán), Argentina. Facena 7: 125-143
Malanca, S., Martínez, O.G.,
Aris, M.J., Boso, M.A., Gallardo, E., Brandán, E. y
Fernández, J.C. 2008. Las plantas fósiles de la Formación
Lipeón (Silúrico Superior) del área Bermejo-Los Toldos,
noroeste argentino. XVII Congreso Geológico Argentino,
Jujuy. Actas: 363
Manca, N., Heredia, S.,
Hunicken, M. y Rubinstein, C. 1995. Macrofauna, conodontes y
acritarcos de la Formación Santa Rosita (Tremadociano),
Nazareno, provincia de Salta, Argentina. Boletín de la
Academia Nacional de Ciencias de Córdoba. 60: 267-275
Martínez, L.C.A. y Lutz, A.I.
2006. Un nuevo género de Fabaceae fósil de la Formación
Chiquimil (Mioceno Superior), Catamarca, Argentina. 9º
Congreso Argentino de Paleontología y Bioestratigrafía,
Córdoba. Resúmenes: 44
Menéndez, C.A. 1962. Leño
petrificado de una Leguminosa del Terciario de Tiopunco,
provincia de Tucumán. Ameghiniana, 2: 121-126
O'Donell, C. 1937. Troncos y
ramas fósiles de dicotiledóneasen el Araucaniano de Tiopunco
(Depto. Tafí, prov. Tucumán). Cuadernos de Mineralogía y
Geología (Universidad Nacional de Tucumán), 1: 26-29
Peirano, A. 1943. Algunos
yacimientos de fósiles de la parte central del valle de
santa María y valle del Cajón. Cuadernos de Mineralogía y
Geología III nº 9: 40-54
Quattrocchio, M.E. 1978b.
Datos paleoecológicos y paleoclimatológicos de la Formación
Lumbrera (Grupo Salta). Ameghiniana, 15: 173-181
Ver mas
bibliografía utilizada para hacer la presente Pagina; AQUÍ.
Ver bibliografía de divulgación sugerida; AQUÍ.
Que información deseas encontrar
en Grupo Paleo?
Nuestro sitio Web posee una amplia
y completa información sobre geología, paleontología,
biología y ciencias afines. Antes de realizar una
consulta por e-mail sobre algunos de estos temas,
utilice nuestro buscador interno. Para ello utilice
palabras "claves", y se desplegara una lista de
"coincidencias". En caso de no llegar a satisfacer sus
inquietudes, comuníquese a
grupopaleo@gmail.com
Queres agregar algo o notificar un error? -
PaleoArgentina es un lugar abierto a toda la comunidad
científica, técnica y aficionados.
grupopaleo@gmail.com












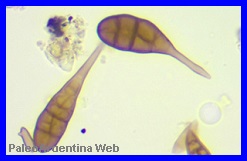



































.jpg)
.JPG)
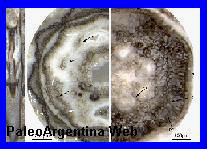















.JPG)
.jpg)
.jpg)

























.jpg)
.jpg)